The genus Streptomyces is a unique subgroup of actinomycetes bacteria that is well-known as prolific products of many bioactive secondary metabolites as well as antibiotics. Streptomyces globosus (S. globosus) is a bacterium species from the genus of Streptomyces. Objective: This study aimed to optimize the growth conditions required for the maximum antibacterial activity of S. globosus isolated from soil samples in Saudi Arabia. The effect of dark/light, temperature, and pH on the growth and antibacterial effect against E. coli ATCC35218, E. coli ATCC25922, Salmonella sp., P. aeruginosa ATCC27853, S. aureus ATCCBAA977, and St. pneumonia ATCC49619 were optimized. The results of this study showed that. The highest antibacterial activity was at 30 ° C after 7 days of incubation.
Furthermore, the best to produce antibacterial activity was under alkaline conditions (pH 8.5). Besides, the isolate didn't show antibacterial activity at the shaking incubator. This study showed that all organic solvents (ethyl-alcohol, methanol, chloroform, ethyl-acetate, and acetone) successfully extracted the antibacterial compound. Still, the compounds extracted by chloroform had the highest antibacterial activity. The MIC of the chloroform extract was against S. aureus ATCCBAA977. The study described the optimized condition for best S. globosus activity. The S. globosus showed a broad spectrum of antimicrobial activities against the test organisms. This opened further research investigations on the purification and structural characterization of the active compounds from the crude extract.
INTRODUCTION
Antimicrobial is used to spare people from contamination. It is a substance or a synthetic item with an inhibitory or destructive impact on the microorganisms (Piddock, 2012; Aloqayli et al., 2022). The advancement of safe strains is a characteristic wonder that happens through choice weight on the antimicrobial microorganism populace (Chellat et al., 2016). Nonetheless, some of these antimicrobials have been rendered dormant by resistant systems (Nikolaidis et al., 2014; Batarseh et al., 2022). Several natural antimicrobials were extracted from soil microbes (Jain & Pundir, 2011; Qinn et al., 2020).
Actinomycetes are a group of prokaryotic microorganisms, gram-positive bacteria with high guanine+cytosine in their DNA (Adegboye & Babalola, 2012). They are considered biotechnologically important organisms since they are responsible for producing about half of the bioactive secondary metabolites, including antibiotics. They are filamentous bacteria that produce two types of branching mycelium, namely aerial and substrate mycelium (Yuan et al., 2010).
Streptomyces (family Streptomycetaceae) are the most significant Actinobacteria (Rong et al., 2009; Radwan et al., 2022). It is disseminated in soil, water, and other indigenous habitats (Singh et al., 2006). In all years, Streptomyces strains as a wellspring of various bioactive mixes, for example, anthelminthic chemicals, herbicides, hostile to malignancy drugs, development components, and insusceptible modulators (Kumar et al., 2020; Adeyemo et al., 2021).
Factors influencing the number and types of actinomycetes present in a particular soil are geographical location, such as soil type, temperature, organic matter content, moisture content, cultivation, and aeration. Actinomycetes act as a major component of the microbial population in most soil. About 90% of the total actinomycetes population consists of Streptomyces species (Harir et al., 2018). Almost 80% of the world's antibiotics are known to come from actinomycetes, mostly from the genera Streptomyces and Micromonospora (Pandey et al., 2004).
A major global healthcare problem has occurred in the 21st century because of the increasing resistance of microorganisms against commonly used antibiotics. A renewed search for new antimicrobial agents from Streptomyces has occurred because of this rapid emergence of antimicrobial resistance among pathogenic microorganisms (Alanis, 2005; Sebak et al. 2021).
Microscopic organisms are profoundly versatile pathogens that create protection from antimicrobials through a few mechanisms. The creation of β-lactamases that hydrolyze the β-lactam ring is the most widely recognized resistant system for these microscopic organisms against b-lactam antimicrobials (Pfeifer et al., 2010; Tooke et al., 2019). Creating board range β-lactamases (ESBLs) is a noteworthy opposition instrument that obstructs the antimicrobial treatment of contaminations brought about by Enterobacteriaceae and is a genuine risk to the presently accessible antimicrobial arsenal (Shaikh et al., 2015). Bacterial efflux pumps effectively transport numerous antimicrobials out of the cell. They are significant contributors to the characteristic resistance of gram-negative microscopic organisms to many medications that can be utilized to treat gram-positive bacterial contaminations (Blair et al., 2015).
This study aimed to optimize the growth conditions required for the maximum antibacterial efficacy of Streptomyces globosus (S. globosus) separated from soil samples in Saudi Arabia.
MATERIALS AND METHODS
Effect of different pH values
The 100 ml Calibration of sterile starch-nitrate broth medium for different pH levels; 6, 6.5, 7, 7.5, 8, 8.5, 9, and 9.5 using Na OH for alkaline and HCl for acidic before sterilizing medium was performed. The cultures were incubated at 30Co. The culture filtrates were centrifuged (4000 rpm in 30 m). The antibacterial efficacy of CFF was investigated using the agar well diffusion technique. Twenty-four hours post-incubation (35 ± 2°C), the antibacterial efficacy was calculated by observing the zone of inhibition radius in millimeters.
Effects of the light factor
Nine Petri plates by the selected streptomyces isolate were incubated (three in light, dark, or 12 h light/ 12 h dark) at 28°C for 7 days. Two agar discs (6 diameters) of Streptomyces isolate were placed over the surface of the tested bacteria's cultures with equal distance and incubated at 35 ± 2 °C for one day. After incubation, the antibacterial activity was estimated by measuring the inhibition zones' diameter in (mm).
Effect of different incubation periods at different incubation temperatures
The chosen Streptomyces sp. was cultured on sterile starch-nitrate broth (100 ml) medium at different temperatures (25°, 30°, 35° and 40° C) for various periods (1, 2, 3, 4, 5, 6,7, 8 and 9). At each incubation period and different temperatures, the culture filtrates were centrifuged (4000 rpm in 30 m). The antibacterial efficacy of cell-free filtrate (CFF) was investigated using the agar well diffusion technique. Twenty-four hours post-incubation (35 ± 2 ° C), the antibacterial efficacy was calculated by observing the zone of inhibition radius in millimeters.
Effect of shaking incubator
The chosen Streptomyces sp. was cultured on sterile starch-nitrate broth (100 ml) medium in optimum conditions. Half of the cultures were incubated on a shaking incubator, and others were incubated in a constant incubator. The culture filtrates were centrifuged (4000 rpm in 30 m). The antibacterial efficacy of CFF was investigated using the agar well diffusion technique. Twenty-four hours post-incubation (35 ± 2 ° C), the antibacterial efficacy was calculated by observing the zone of inhibition radius in millimeters.
Extraction of an antibacterial compound with organic solvents
Four different solvents (ethyl-alcohol, methanol, chloroform, ethyl-acetate, and acetone) were used to extract the antibacterial activity agent that was produced by the selected Streptomyces sp. The chosen Streptomyces sp. was cultured on sterile starch-nitrate broth (100 ml) medium in optimum conditions. The culture filtrates were centrifuged (4000 rpm in 30 m). The CFF was transferred aseptically into the conical flasks, and an equal volume of solvents was separately added to the CFF and shaken for 20 min. After gathering the organic phase, the solvent was evaporated under a vacuum. 50 µl of the extract was used to investigate antibacterial efficacy using the agar well diffusion technique. Twenty-four hours post-incubation (37 ° C), the antibacterial efficacy was calculated by observing the zone of inhibition radius in millimeters.
MIC values against nosocomial infection bacteria
The process includes the preparation of a double dilution of the antimicrobial agents in a liquid medium. Upon diluting the standardized bacterial culture, each tube with a bacterial suspension was packed in the same medium. The inoculated tubes were incubated (mostly without agitation) under suitable conditions depending upon the test microorganism.
Statistics
The data presented as a mean of 3 replicates ± SD. The difference between the means was statistically compared using Microsoft Excel software.
rESULTS AND DISCUSSION
Effect of the light factor on antibacterial activity of S. globosus
Table 1 revealed no significant difference in the antibacterial activity when S. globosus was grown in different lighting. It had antibacterial activity against all the test bacteria except P.earuginosa ATCC27853, but no differences in the antibacterial activity against the other test bacterial isolates when S. globosus was grown in light, dark, or 12h light / 12h dark, it showed antibacterial activity against E.coli ATCC35218 only when it was grown in dark.
Table 1. Effect of the light factor on antibacterial activity of S. globosus
|
Light factors |
Inhibition zone diameter (mm) |
|||||
|
E. coli ATCC35218 |
E. coli ATCC25922 |
Salmonella sp. |
P. earuginosa ATCC27853 |
S. aureus ATCCBAA977 |
St. pneumonia ATCC49619 |
|
|
Dark |
11.3 ± 0.47 |
15.6 ± 0.47 |
18.3 ± 0.47 |
- |
24.3 ± 0.47 |
14.6 ± 0.47 |
|
Light |
- |
15.3 ± 0.47 |
17.16 ± 0.23 |
- |
23.16 ± 0.23 |
13.3 ± 0.23 |
|
12h light / 12h dark |
- |
15.3 ± 0.47 |
17.83 ± 0.23 |
- |
24 ± 0.00 |
14.3 ± 0.47 |
Effect of different incubation periods and different incubation temperatures on antibacterial activity of S. globosus
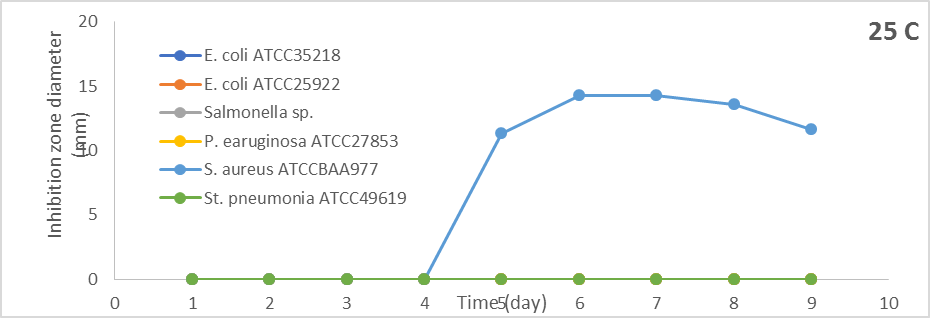
Incubation of S. globosus at 25oC showed antibacterial activity against S. aureus ATCCBAA977 only after five days of incubation and increased by increasing the incubation period. The most excellent antibacterial activity was after seven days of incubation, and the inhibition zone was 14.3 mm. Still, there was no antibacterial activity against other Gram-positive and Gram-negative bacteria (Figure 1).
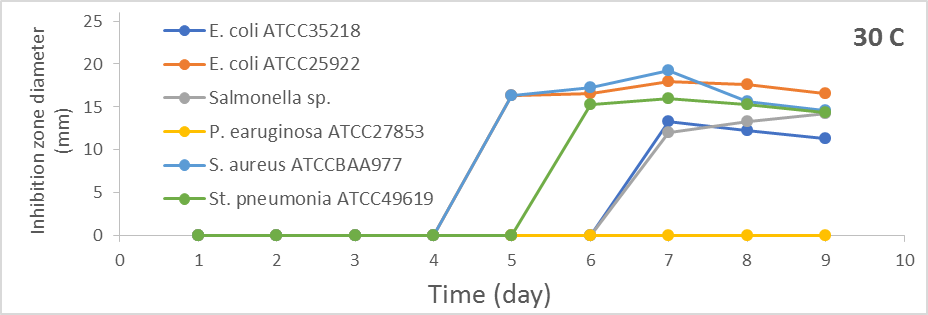
Results in Figures 1a-1c revealed that antibacterial activity against most test bacteria was noticed and increased with increasing the incubation period at 30oC. The optimum antibacterial activity was after seven days of incubation. The diameters of inhibition zones were (13.3 mm, 18.0 mm, 12.0 mm, 19.3 mm, and 16.3 mm) against E. coli ATCC35218, E. coli ATCC25922, Salmonella sp., S. aureus ATCCBAA977 and St. pneumonia ATCC49619, respectively.
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
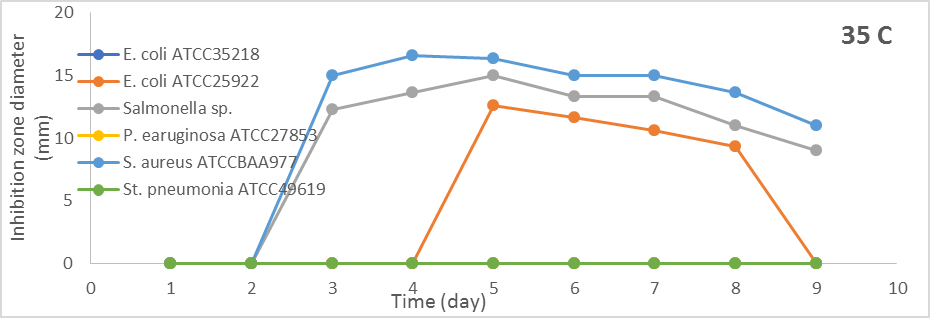
Figure 1. Effect of different incubation periods and different incubation temperatures on antibacterial activity of S. globosus |
From the results in Figure 1, when S. globosus was incubated at 35oC, the antibacterial efficacy was versus E. coli ATCC25922, Salmonella sp., and S. aureus ATCCBAA977. The antibacterial activity against Salmonella sp. and S. aureus ATCCBAA977 began from the third day but after five days produced antibacterial activity against E. coli ATCC25922. The optimum antibacterial activity was on the fifth day, and the diameters of inhibition zones were (12.6 mm, 15.0 mm, and 16.3 mm) versus E. coli ATCC25922, Salmonella sp, and S. aureus ATCCBAA977. S. globosus at 40oC did not show antibacterial activity against all test bacteria.
Impact of various pH conditions on antibacterial efficacy of S. globosus
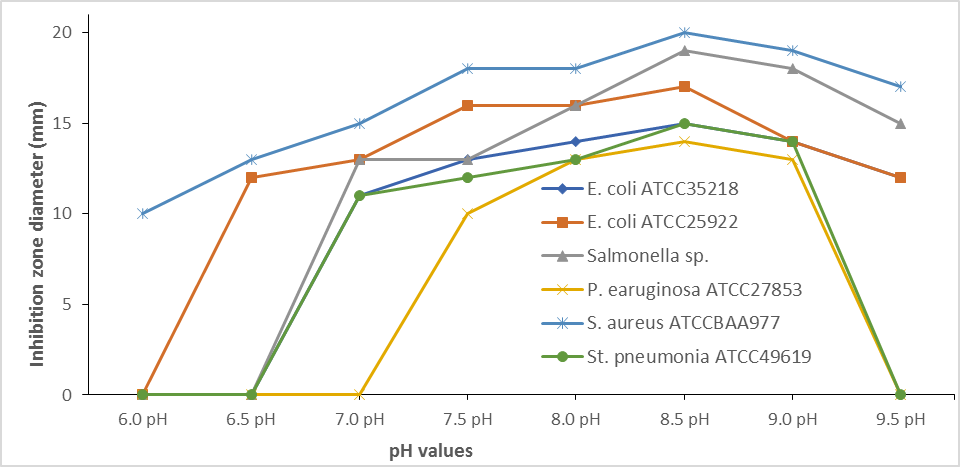
S. globosus produce antibacterial activity against S. aureus ATCCBAA977 at pH 6, against E. coli ATCC25922, and S. aureus ATCCBAA977 at pH 6.5, and against all test bacterial isolate except P. earuginosa ATCC27853 at pH 7. Figure 2 shows that the antibacterial activity of S. globosus against all test bacteria increased by increasing the fermentation medium's pH values. The highest antibacterial activity was at pH 8.5, which was decreased above. It was noticed that S. globosus produced antibacterial activity against all test isolates and the highest was against S. aureus ATCCBAA977. The inhibition zone diameter was (20.3 mm) and decreased slowly above.
|
|
|
Figure 2. Impact of various pH efficacy of S. globosus. condition on antibacterial |
Effect of shaking incubator on antibacterial activity of S. globosus
Results in Figure 3 show the difference between the antibacterial activity produced by S. globosus when the cultures of this isolate were incubated in a constant and shaking incubator. When incubated in a shaking incubator, S. globosus can't have antibacterial activity agents against all the test bacteria. Different antibacterial activities were produced when cultures of S. globosus were incubated in a constant incubator.
|
|
|
Figure 3. Effect of shaking incubator on antibacterial activity of S. globosus. |
The diameters of inhibition zones were (15.3 mm, 14.0 mm, 13.3 mm, 14.3 mm, 24.0 mm, and 15.3 mm) against E. coli ATCC35218, E. coli ATCC25922, Salmonella sp., P. earuginosa ATCC27853, S. aureus ATCCBAA977 and St. pneumonia ATCC49619, respectively.
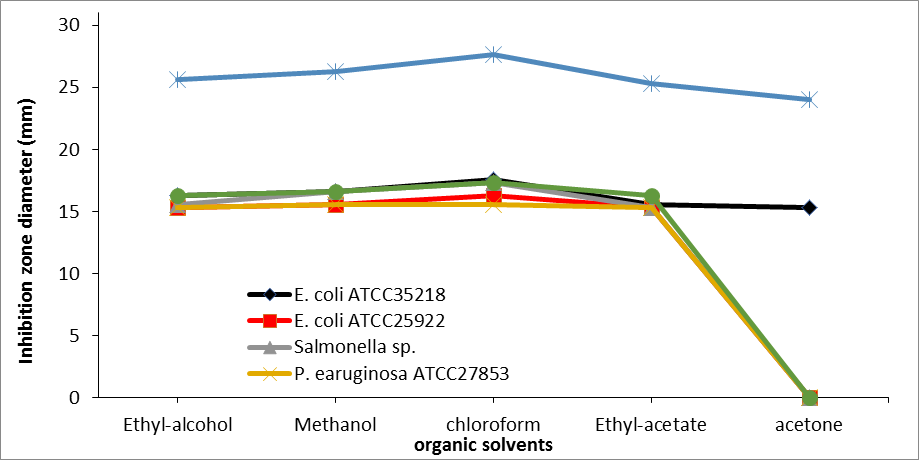
Effect of different organic solvents used in the extraction of the antibacterial compound from S. globosus on its antibacterial activity.
Figure 4 revealed that all organic solvents succeeded in extracting the antibacterial compound. Better extraction was obtained using chloroform. The diameters of inhibition zones were (17.6 mm, 16.3 mm, 17.3 mm, 15.6 mm, 27.6 mm, and 17.3 mm) against E. coli ATCC35218, E. coli ATCC25922, Salmonella sp., P. earuginosa ATCC27853, S. aureus ATCCBAA977 and St. pneumonia ATCC49619, respectively.
|
|
|
Figure 4. Effect of different organic solvents used to extract the antibacterial compound from S. globosus on its antibacterial activity. |
MIC values of the chloroform extract against nosocomial infection bacteria
The MIC values against all the tested bacteria showed different results. S. aureus ATCCBAA977 has been inhibited at (16 µg/m), which was the lowest concentration for inhibition. E. coli ATCC35218, Salmonella sp., and St. Pneumonia ATCC49619 bacteria have been inhibited at (32 µg/ml). E. coli ATCC25922 inhibited at (64 µg/ml). The highest concentration for inhibition was at (125 µg/ml) against P. earuginosa ATCC27853 (Table 2).
Table 2. MIC values of the chloroform extract against nosocomial infection bacteria.
|
Nosocomial infection bacteria |
Minimum inhibitory concentration (MIC) |
|||||
|
E. coli ATCC35218 |
E. coli ATCC25922 |
Salmonella sp. |
P. earuginosa ATCC27853 |
S. aureus ATCCBAA977 |
St. pneumonia ATCC49619 |
|
|
Chloroform |
32 |
64 |
32 |
125 |
16 |
32 |
In this work, Streptomyces sp. was dominantly separated from the soil specimen on a starch-nitrate medium. This finding is congruent with earlier studies, which reported that the recovery percentage of Streptomyces sp. with antimicrobial activity was higher than other actinobacterial genera (Kitouni et al., 2005). Rare actinobacteria are also important in discovering novel antibiotics (Tiwari & Gupta, 2012).
This study aimed to optimize the culture condition, which will yield the most optimum antimicrobial efficacy of Streptomyces sp. The incubation temperature can influence bacterial growth and the development of biosynthetic pathways of the metabolites (Stanbury, 2016). Also, Pandey et al. (2005) and Vijayakumar et al. (2012) showed that for the optimum production of antibiotics, certain carbon and nitrogen sources are required study done by Vasavada et al. (2006) showed that the use of different media, pH, salinity, and carbon and nitrogen affect the growth and antibiotic production by Actinomycetes.
Results of the impact of incubation time and temperature on the optimum efficacy of S. globosus showed that the highest antibacterial activity was at 30 ° C after 7 days of incubation. This result is similar to Sujatha et al. (2005), with the highest antimicrobial activity at 30 ° C. On the other hand, Ripa et al. (2009) reported that isolated Streptomyces sp. produced high antibiotic levels when incubated at 39 °C.
This study showed that the best to produce antibacterial activity was under alkaline conditions (pH 8.5). Similar results had been reported earlier by Basilio et al. (2003). Ripa et al. (2009) showed that the pH 8.0 was optimum for antibiotics produced by Streptomyces sp. This may be attributed to the presence of active enzymes for antimicrobial metabolites synthesis at pH 7-9. Each Streptomyces strain has an optimum, minimum, and maximum pH at which it gave an optimum enzyme activity (Vijayakumar et al., 2012).
This study showed that the isolate didn't show antibacterial activity at the shaking incubator. Gottschalk et al. (2003), in their research using S. viridosporus, suggested that the rise in the rate of agitation up to 500 rpm contributed to a persistent decline in the growth and development of cells. A rise in the stirring rate may have contributed to high shearing outcomes that were harmful to the solidity of the fungi.
This study showed that all organic solvents (ethyl-alcohol, methanol, chloroform, ethyl-acetate, and acetone) successfully extracted the antibacterial compound. Still, the compounds extracted by chloroform had the highest antibacterial activity. Our results were similar to those of Sathi et al. (2001). Disagree with our results, Vijayakumar et al. (2012) confirmed that extraction of Streptomyces sp. with ethyl acetate produced more activity by generating the maximum inhibition versus P. Vulgaris and minimum inhibition versus S. aureus. Besides, Balachandran et al. )2012) showed that extraction of Methylobacterium sp. With ethyl acetate produced the maximum inhibition versus K. pneumonia and minimum inhibition versus B. subtilis.
CONCLUSION
These findings indicated that the soil samples of the Al-Baha region have actinomycetes with metabolites that control bacterial pathogens. The extracted metabolites obtained from the isolate of our study showed very good antimicrobial activity against four different clinical pathogens. The further structural characterization of these molecules promotes the utilization of this active principle as a lead in case of drug discovery from soil-based formulations to eradicate the emerging infectious drug-resistant pathogenic microorganisms.
ACKNOWLEDGMENTS: None
CONFLICT OF INTEREST: None
FINANCIAL SUPPORT: None
ETHICS STATEMENT: None
Adegboye, M. F., & Babalola, O. O. (2012). Taxonomy and ecology of antibiotic producing actinomycetes. African Journal of Agricultural Research, 7(15), 2255-2261.
Adeyemo, O. M., Ja’afaru, M. I., & Adams, F. V. (2021). Isolation, characterization, antimicrobial and other bioactivity profiles of three Streptomyces strains isolated from Lake Gerio, Yola, Adamawa State, Nigeria. Bulletin of the National Research Centre, 45(1), 1-13. doi:10.1186/s42269-021-00606-x
Alanis, A. J. (2005). Resistance to antibiotics: Are we in the post-antibiotic era? Archives of Medical Research, 36(6), 697-705.
Aloqayli, M. A., Alshuwirikh, R. M., Alghwery, N. S., Ansari, S. H., & Alhussain, B. S. (2022). Quality assessment of composite restorations performed by the dental students: A retrospective study. Annals of Dental Specialty, 10(2), 101-105.
Balachandran, C., Duraipandiyan, V., Al-Dhabi, N. A., Balakrishna, K., Kalia, N. P., Rajput, V. S., Khan, I. A., & Ignacimuthu, S. (2012). Antimicrobial and antimycobacterial activities of methyl caffeate isolated from Solanum torvum Swartz. fruit. Indian Journal of Microbiology, 52(4), 676-681.
Basilio, A., Gonzalez, I., Vicente, M. F., Gorrochategui, J., Cabello, A., Gonzalez, A., & Genilloud, O. (2003). Patterns of antimicrobial activities from soil actinomycetes isolated under different conditions of pH and salinity. Journal of Applied Microbiology, 95(4), 814-823.
Batarseh, N., Khalil, R., & Al-Domi, H. A. (2022). Hypothalamic neuroinflammation induced by obesity and the effect of Liraglutide. Journal of Advanced Pharmacy Education and Research, 12(1), 46-55.
Blair, J., Webber, M. A., Baylay, A. J., Ogbolu, D. O., & Piddock, L. J. (2015). Molecular mechanisms of antibiotic resistance. Nature Reviews Microbiology, 13(1), 42-51.
Chellat, M. F., Raguž, L., & Riedl, R. (2016). Targeting antibiotic resistance. Angewandte Chemie International Edition, 55(23), 6600-6626.
Gottschalk, L. M., Nobrega, R., & Bon, E. P. (2003). Effect of aeration on lignin peroxidase production by Streptomyces viridosporus T7A. In Biotechnology for Fuels and Chemicals (pp. 799-807). Humana Press, Totowa, NJ.
Harir, M., Bendif, H., Bellahcene, M., Fortas, Z., & Pogni, R. (2018). Streptomyces secondary metabolites. Basic Biology and Applications of Actinobacteria, 6, 99-122.
Jain, P., & Pundir, R. K. (2011). Effect of fermentation medium, pH and temperature variations on antibacterial soil fungal metabolite production. Journal of Agricultural Technology, 7(2), 247-269.
Kitouni, M., Boudemagh, A., Oulmi, L., Reghioua, S., Boughachiche, F., Zerizer, H., Hamdiken, H., Couble, A., Mouniee, D., Boulahrouf, A., et al. (2005). Isolation of actinomycetes producing bioactive substances from water, soil and tree bark samples of the north–east of Algeria. Journal de Mycologie Médicale, 15(1), 45-51.
Kumar, P. S., Ling, C. Y., Zhou, Z. B., Dong, Y. L., Sun, C. L., Song, Y. X., Wong, N. K., & Ju, J. H. (2020). Chemical diversity of metabolites and antibacterial potential of actinomycetes associated with marine invertebrates from intertidal regions of Daya Bay and Nansha Islands. Microbiology, 89(4), 483-492.
Nikolaidis, I., Favini‐Stabile, S., & Dessen, A. (2014). Resistance to antibiotics targeted to the bacterial cell wall. Protein Science, 23(3), 243-259.
Pandey, A., Shukla, A. N. U. P. A. M., & Majumdar, S. K. (2005). Utilization of carbon and nitrogen sources by Streptomyces kanamyceticus M 27 for the production of an Anti bacterial antibiotic. African Journal of Biotechnology, 4(9), 909-910.
Pandey, B., Ghimire, P., & Agrawal, V. P. (2004). International Conference on the Great Himala-yas: Climate, Health, Ecology, Management and Conservation, Kathmandu.
Pfeifer, Y., Cullik, A., & Witte, W. (2010). Resistance to cephalosporins and carbapenems in Gram-negative bacterial pathogens. International Journal of Medical Microbiology, 300(6), 371-379.
Piddock, L. J. (2012). The crisis of no new antibiotics—What is the way forward? The Lancet Infectious Diseases, 12(3), 249-253.
Quinn, G. A., Banat, A. M., Abdelhameed, A. M., & Banat, I. M. (2020). Streptomyces from traditional medicine: Sources of new innovations in antibiotic discovery. Journal of Medical Microbiology, 69(8), 1040-1048.
Radwan, W. W., AlTuwaijri, D. S., Alwoseamer, A. T., & AlMajed, A. I. (2022). Endodontically treated teeth restoration assessment, decision making and treatment option among dental practitioners in Saudi Arabia. Annals of Dental Specialty, 10(1), 118-125.
Ripa, E. A., Nikkon, K., Zaman, S., & Khondkar, P. (2009). Optimal conditions for antimicrobial metabolites production from a new Streptomyces sp. RUPA-08PR isolated from Bangladeshi soil. Mycobiology, 37(3), 211-214.
Rong, X., Guo, Y., & Huang, Y. (2009). Proposal to reclassify the Streptomyces albidoflavus clade on the basis of multilocus sequence analysis and DNA–DNA hybridization, and taxonomic elucidation of streptomyces griseus subsp. solvifaciens. Systematic and Applied Microbiology, 32(5), 314-322.
Sathi, Z. S., Rahman, M. A. A., & Gafur, M. A. (2001). Identification and in vitro antimicrobial activity of a compound isolated from streptomyces species. Pakistan Journal of Biological Sciences, 4, 1523-1525.
Sebak, M., Saafan, A. E., Abdelghani, S., Bakeer, W., Moawad, A. S., & El-Gendy, A. O. (2021). Isolation and optimized production of putative antimicrobial compounds from Egyptian soil isolate streptomyces sp. MS. 10. Beni-Suef University Journal of Basic and Applied Sciences, 10(1), 1-12.
Shaikh, S., Fatima, J., Shakil, S., Rizvi, S. M. D., & Kamal, M. A. (2015). Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi Journal of Biological Sciences, 22(1), 90-101.
Singh, L. S., Baruah, I., & Bora, T. C. (2006). Actinomycetes of Loktak habitat: Isolation and screening for antimicrobial activities. Biotechnology, 5(2), 217-221.
Stanbury, P. F. (2016). Principles of Fermentation Technology Ed. 3. Elsevier Science.
Sujatha, P., Raju, K. B., & Ramana, T. (2005). Studies on a new marine streptomycete BT-408 producing polyketide antibiotic SBR-22 effective against methicillin resistant Staphylococcus aureus. Microbiological Research, 160(2), 119-126.
Tiwari, K., & Gupta, R. K. (2012). Rare actinomycetes: A potential storehouse for novel antibiotics. Critical Reviews in Biotechnology, 32(2), 108-132.
Tooke, C. L., Hinchliffe, P., Bragginton, E. C., Colenso, C. K., Hirvonen, V. H., Takebayashi, Y., & Spencer, J. (2019). β-Lactamases and β-Lactamase inhibitors in the 21st century. Journal of Molecular Biology, 431(18), 3472-3500. doi:10.1016/j.jmb.2019.04.002
Vasavada, S. H., Thumar, J. T., & Singh, S. P. (2006). Secretion of a potent antibiotic by salt-tolerant and alkaliphilic actinomycete streptomyces sannanensis strain RJT-1. Current Science, 1393-1397.
Vijayakumar, R., Panneerselvam, K., Muthukumar, C., Thajuddin, N., Panneerselvam, A., & Saravanamuthu, R. (2012). Optimization of antimicrobial production by a marine actinomycete streptomyces afghaniensis VPTS3-1 isolated from palk strait, east coast of India. Indian Journal of Microbiology, 52(2), 230-239.
Yuan, L. J., Zhang, Y. Q., Yu, L. Y., Sun, C. H., Wei, Y. Z., Liu, H. Y., Li, W. J., & Zhang, Y. Q. (2010). Actinopolymorpha cephalotaxi sp. nov., a novel actinomycete isolated from rhizosphere soil of the plant cephalotaxus fortunei. International Journal of Systematic and Evolutionary Microbiology, 60(1), 51-54.
 This work is licensed under a Creative Commons Attribution 4.0 International License.
This work is licensed under a Creative Commons Attribution 4.0 International License.