The ever-increasing predicament of climate change results in major alterations in the immediate environment of different organisms at an unprecedented global scale. Thus, organisms find themselves trapped in an environment that alters at a rate much higher than their genotypes can adapt, owing to their relaxed rates of mutation. This is exactly where epigenetic modifications come in handy, as they help the organism cope with drastic changes in environmental conditions mediated by climate change. That said, these adaptations may not be enough to combat the rapidly-changing climate changes in the long run. Furthermore epigenetic modification-mediated adaptations are not always for the greater good; in certain cases, it may result in the organism becoming invasive, in which case other organisms which occupy the same niche, are also affected. Not all epigenetic modifications can be credited with enhanced adaptability in different organisms, and in certain cases have even been proven to be deleterious to the organism itself so much so that it threatens their survival on this planet. Along with climate change, environmental toxicants which are also on the rise lead to detrimental epigenetic alterations that ultimately manifest as life-threatening disease pathologies across different organism lineages.
INTRODUCTION
Unlike ‘genetics’ which involves the study of the entire genome of an organism, ‘epigenetics’ (“epi” means “above”) stands for ‘over and above genetics’. In other words, epigenetics determines the location (for example tissue or cell-specific) and timing of gene expression, in response to the influence of non-genetic factors such as environment, behavior, and lifestyle patterns. Thus, while genetic changes determine which protein will get translated from a particular gene, epigenetic changes regulate the state of the genes which are to be transcribed and translated, based on non-genetic factors (Morgan, 2019).
Studies have shown that in mammals, potent epigenetic changes occur while the cells are differentiating, thus initiating cell-type-specific programs. These epigenetic changes manifest themselves as covalent modifications, primarily in the form of post-synthetic modifications of DNA, RNA, and proteins (Carneiro & Lyko, 2020).
Epigenetic changes at the molecular level are mainly brought about by histone and non-histone proteins. The N-terminal tails of the histone proteins play a pivotal role in epigenetic regulations since they are capable of harboring modifications brought about by different enzymes, which can incorporate and strike out such modifications, depending on the regulatory mechanism. Such modifications are post-translational and possess the ability to alter the chromatin structure (Carneiro & Lyko, 2020). Apart from the above-mentioned mechanisms, several mechanisms based on RNA molecules, such as siRNAs (small interfering RNAs) and miRNAs (microRNAs) have been shown to participate in epigenetic regulations, as they are capable of mediating the degradation of mRNA or hindering its translation (Carneiro & Lyko, 2020). Another possible mechanism by which siRNA carries out epigenetic regulation is via suppression of transposable genetic elements, along with participation in various cross-talks with DNA methylation and additional mechanisms like histone modification (Carneiro & Lyko, 2020).
Long-term increments in global temperature and altered weather patterns are the manifestations of climate change (https://www.un.org/en/climatechange/what-is-climate change). While climate change affects the entire planet, epigenetics involves changes in gene expression in individual cells. What unites these two concepts is the ‘environment’. Contrary to natural climate changes, the climate change that our planet is currently experiencing is mainly anthropogenic, irreversible, and appears to be permanent.
An increase in global temperatures leads to a multitude of alterations in the environment; such changes include, but are not limited to, intense heat waves, altered rainfall patterns, rising sea levels due to the melting of polar ice sheets, changes in salinity, as well as pH of oceans and other water-bodies (United Nations, 2012).
Needless to say, such changes adversely affect global ecosystems due to alterations in the abiotic factors, which in turn, result in changes to the biotic factors i.e., the organisms which inhabit the said ecosystems. The epigenome of an organism encompasses all factors, components, and compounds that are capable of interacting with the genome and modulating its function. Such epigenomes can detect changes in the environment of the organism, and cause changes in the gene expression in its cells. This is an adaptive strategy employed by organisms to get acclimated to environments at a quicker pace, relative to changes in the genome which require a long time to change due to the slow rate of mutations, thereby preventing the organism from becoming extinct.
Recent reports have pointed out that climate change can uninterruptedly manipulate ecospheres. Analysis of epigenomes of the organisms which inhabited such ecospheres has shown that the changes occurring as a result of climate change can be tracked down to their epigenomes themselves, which underwent considerable modifications themselves (Dixon et al., 2018).
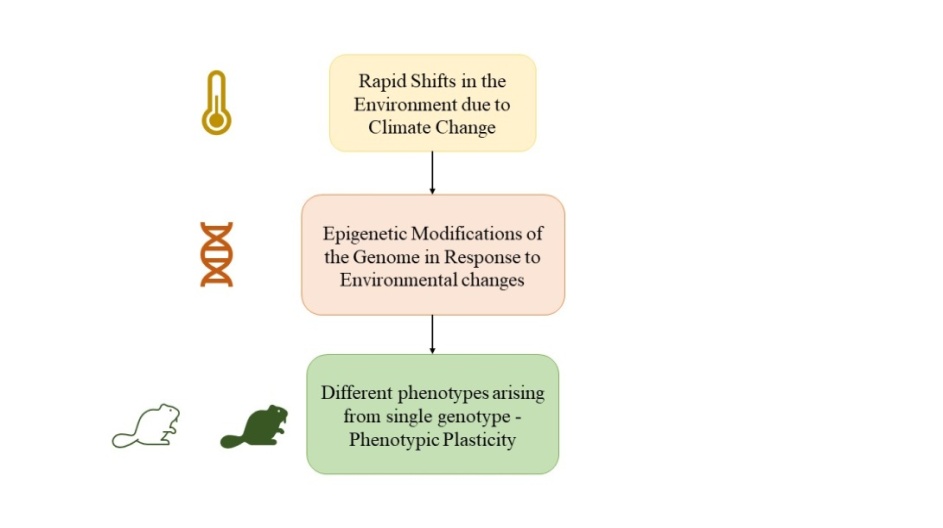
Also, the use of the terms ‘climate change’ and ‘epigenetics’ has increased so much over the last few years that “plasticity” has emerged as an important parameter (Kingsolver & Buckley, 2017). ‘Plasticity’ refers to the extent to which the organisms (plant or animal) can react more coherently to climate- or environment-related irritants as permitted by their epigenomes (Kingsolver & Buckley, 2017). An organism can react to different environmental conditions differently by manifesting the same genotype in the form of different phenotypes (Bonamour et al., 2019). This mechanism of ‘phenotypic plasticity’ is instrumental in tackling dramatic environmental changes as a result of climate change (Figure 1) (Bonamour et al., 2019). A key aspect in this regard is the heritability of the said ‘plasticity’ i.e. whether the epigenetic changes that are brought about by climate change are transmitted to the next generation, resulting in transgenerational plasticity (Morgan, 2019).
|
|
|
Figure 1. Schematic showing the role of epigenetic modifications in ‘Phenotype Plasticity’ |
The fact that the epigenome of an organism can change in response to changes in the environment makes it a very important aspect to study and consider when we try to determine the effects that the major changes in environments brought about by climate change have on the organisms. Temperature is one of the most important factors in this regard, affecting both terrestrials as well as aquatic ecosystems (Morgan, 2019). High temperatures lead to drought on land, while the increase in the temperature of water bodies leads to alteration in salinity and pH, in turn affecting marine life forms (Morgan, 2019). Plants, which are the most affected by these drought stressors due to their inability to move out, are important samples to study epigenetic responses (Müller & Gailing, 2019).
In this review, we will discuss how climate change, environmental toxicants, and epigenetics relate to one another and also shed some light on the adaptive value of epigenetics in the face of climate change.
Climate-change-mediated adaptive epigenetic changes
Climate change has resulted in changes in the environment all around the world at unprecedented levels in an extremely short time. The genomes of organisms are unable to keep up with these changes, due to which organisms find themselves lacking the genes to acclimate themselves to the new environment (Bautista & Crespel, 2021). This problem of lack of adaptability is solved by epigenetic modifications which occur within the cells of the organism in response to changes in the environment, allowing organisms to rapidly adjust to the environment and prevent their extinction (Bautista & Crespel, 2021).
Phenotypic plasticity
There are several examples of displays of ‘phenotypic plasticity’ in response to changes in the environment across different organisms. In the case of Anolis sagrei (the brown anole lizard), upon exposing them to a new environment, methylation patterns that differed from the methylation patterns shown by lizards in the original habitat were observed (Carneiro & Lyko, 2020). The genes which exhibited these differential methylation patterns were linked to ‘phenotypic plasticity’ in the lizards (Carneiro & Lyko, 2020). In the case of an aquatic ecosystem, the genome of Stylophorapistillata (scleractinian coral) shows differential methylation patterns due to an increase in temperature, and a decrease in pH of the ocean waters (Liew et al., 2018). These changes lead to a decrease in cryptic transcription of genes which are meant to be silent, and variable gene expression, resulting in the expression of proteins that promote symbiotic interactions with zooxanthellae algae to prevent the algae from getting expelled from the coral, leading to coral bleaching (Carneiro & Lyko, 2020).
Relationship between plant epigenetics and climate change
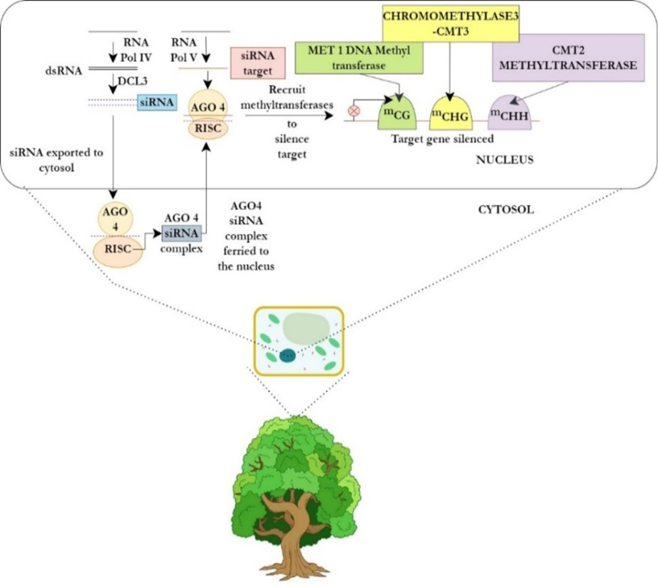
Plants are one of the major terrestrial victims of climate change, owing to their inability to locomote and their long life spans (Müller & Gailing, 2019). DNA methylation is one of the most common mechanisms governing epigenetic regulation in plants, whereby a methyl group is inserted at the position of the fifth carbon within a cytosine ring (Thiebaut et al., 2019). In sharp contrast to DNA methylation in animals, in plants, three positions are very prone to methylation- CHG (where ‘H’ stands for A, T, or C), CG, and CHH (Thiebaut et al., 2019). By performing various experiments, researchers have disclosed the various enzymes that catalyze the process of DNA methylation in plants (Thiebaut et al., 2019). It has been found that separate methyl transferases are responsible for DNA methylation under different circumstances (Thiebaut et al., 2019). For example, while CHG methylation is mediated by Chromomethylase3 (CMT3), CG methylation is taken care of by MET1 DNA Methyl transferase, whereas CHH methylation is maintained by CMT2 Methyltransferase or Domains Rearranged Methyltransferase (DRM)-1 or -2 (Thiebaut et al., 2019) (Figure 2).
|
|
|
Figure 2. The major mechanisms governing epigenetic regulation in plants (Thiebaut et al., 2019) |
Small interfering RNAs (siRNAs), which are procured in the nucleus from the RNA Polymerase IV-mediated transcription of double-stranded (ds)-RNAs and handled by DICER-LIKE 3 (DCL3), also participate in epigenetic regulation in plants by directing the pathway of RNA-directed DNA Methylation (RdDM) (Thiebaut et al., 2019). Following the formation and export of siRNAs to the cytosol, they are loaded onto the Argonaute4 (AGO4)-containing complex RISC (RNA-Induced Silencing Complex) (Thiebaut et al., 2019). The siRNA-AGO4 complex is then ferried to the nucleus where the siRNA orients itself with respect to its target and an RNA Polymerase V-mediated nascent scaffold transcript, DNA methyl transferases get recruited, and the target gets silenced (Thiebaut et al., 2019)(Figure 2).
Abiotic stress can also give rise to altered
DNA methylation patterns in plants (Sow et al., 2018). Such epigenetic modifications provide plants with ‘phenotypic plasticity’, which they can use to overcome the various stressors imposed on them by climate change-mediated environmental changes, such as drought (Sow et al., 2018). This is especially true for forest trees, which depend on epigenetic modification-mediated ‘phenotypic plasticity’ to combat multiple adverse environmental factors, often simultaneously (Sow et al., 2018). DNA methylation has been shown to not only affect biomass but also regulate gene expression, resulting in ‘phenotypic plasticity’ in poplar trees (Le Gac et al., 2018). Variations in soil water content have also been correlated to changes in DNA methylation patterns, especially affecting genes involved in phytohormone production and cell signaling, which contributes to ‘phenotypic plasticity’ (Lafon-Placette et al., 2018).
Epigenetic modifications may not only help plants combat the stress but also help them recover from it, thereby increasing their chances of survival should adverse environmental conditions return (Sow et al., 2018). In the case of conifers (drought-sensitive), it has been hypothesized that the epigenetic mechanisms are involved in the recovery of the trees after exposure to drought; the absence of these mechanisms results in the death of the trees after the drought, as they lacked resilience (Figure 2) (Sow et al., 2018).
Epigenetic modifications and guinea pigs
In a novel experiment performed by Weyrich et al. they not only showed that certain epigenetic modifications may be trans-generational i.e. pass from parent to offspring, but also how specific the epigenetic modification was to the environmental cue that caused it (Weyrich et al., 2018). They also provided a detailed explanation of the adaptive value of the said modification (Weyrich et al., 2018).
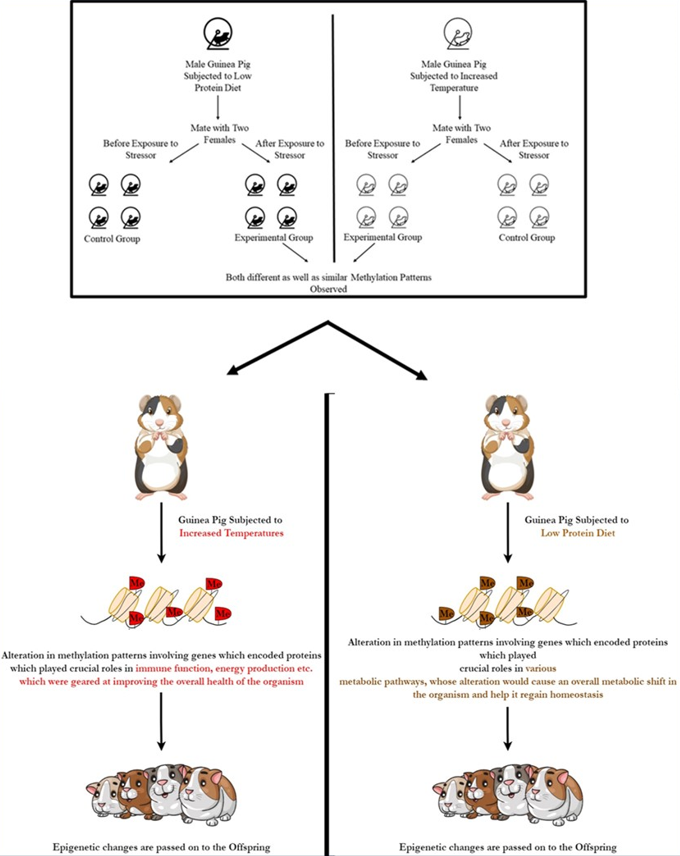
For the experiment, two sets of wild male guinea pigs were taken. One set was exposed to an altered diet which was low in protein, while the other set was subjected to increased temperatures (Figure 3). Each male was made to mate with two females both before and after exposure (same two females). The offspring born as a result of mating before the exposure to altered conditions (either altered diet or temperature) acted as controls in each case. Following this, methylation patterns in the liver of the offspring were studied (Weyrich et al., 2018).
|
|
|
Figure 3. Epigenetic modifications are Stress-specific, General and Trans-genenerational (Weyrich et al., 2018) |
The results showed that the offspring inherited the father’s epigenetic response (Weyrich et al., 2018). They also showed that the offsprings showed some factor-dependent (i.e., some of the epigenetic modifications were different between the two groups of offsprings) as well as certain factor-independent (i.e., some of the epigenetic modifications were similar between the two groups of offspring) epigenetic modifications (Weyrich et al., 2018). Based on this, they hypothesized that epigenetic modifications conferred two types of information to the next generation; firstly, the fact that the environment had changed (conveyed by the factor independent-modifications), also referred to as “alert information”; and, secondly, the information regarding which factor had changed (conveyed by the factor-dependent modifications), also referred to as “adequate response information” (Weyrich et al., 2018). Thus, these modifications act as a means by which parents can prepare their offspring to combat the different environmental stresses to improve their chances of survival (Weyrich et al., 2018).
In addition to this, methylation patterns of the liver showed that an offspring fathered by males exposed to a low protein diet showed altered methylation patterns involving genes that encoded proteins that played crucial roles in various metabolic pathways, so much so that their alteration would cause an overall metabolic shift in the organism and help it regain homeostasis (Weyrich et al., 2018). However, the case of an offspring fathered by males exposed to increased temperatures showed altered methylation patterns involving genes responsible for immune function and energy production which were geared at improving the overall health of the organism (Weyrich et al., 2018). This showed that the epigenetic modifications were not only transgenerational but also adaptive, as well as, to some extent factor-dependent (Weyrich et al., 2018).
Epigenetics-mediated adaptive phenotypic response in winter skates
The hypothesis under investigation was that differential gene expression as a result of different climatic circumstances can lead to the development of considerable variations with respect to the morphology and ecophysiology of populations of wild fishes (Lighten et al., 2016).
It is already known that temperature might be an important parameter that influences various aspects of the fish’s lives, such as growth pattern timings and attributes of life history related to reproduction (Lighten et al., 2016; Rezapour-Nasrabad & Tayyar-Iravanlou, 2022).
For this experiment, two populations of the Skate fishes were collected - one being Winter Skates from the southern Gulf of St. Lawrence (sGSL) population, and the other being the Scotian Shelf population (Lighten et al., 2016). Prior studies have revealed that the sGSL population is endemic and considerably distinct from the Scotian Shelf population with respect to morphology (as it has a remarkably smaller body size), ecophysiology (as it colonizes in markedly warmer waters with temperature differences of up to 10οC) and life span (as they have a brief life span) (Lighten et al., 2016).
Skates possess attributes of a typical K-strategist on account of their long period of maturation, and comparatively low fertility (Lighten et al., 2016). This indicates that they should be less susceptible to drastic evolutionary changes (Lighten et al., 2016). Perversely, it has been observed that skates have unexpected degrees of endemism and diversity across the planet (Lighten et al., 2016). Therefore, in this case, the concept of phenotypic adaptation mediated by epigenetic modifications becomes even more significant as opposed to Fisher’s concept of natural selection-mediated gradual modifications in allelic frequencies (Eoh & Rhee, 2013).
So, to figure out the appropriate hypothesis regarding the diversity of the K-strategist Skates, transcriptomes from both the Skate populations were compared, which revealed the crucial groups of genes in charge of differential attributes with respect to ecophysiology and morphology in both the Skate populations and their variable expression patterns (Lighten et al., 2016). It was observed that the differences in the expression of genes could be traced back to climate change-mediated regional adaptations (Lighten et al., 2016). For example, in the case of the sGSL population, transcriptional down-regulation of genes corresponding to reactions such as hypoxia, X-rays, lower temperatures, and ultraviolet (UV)-rays were observed since they are associated with the subjection of such populations to considerably increased levels of light and decreased aerobic volumes in fishes which reside in waters with elevated temperatures (Cheng et al., 2013). Additionally, significant degrees of down-regulation of transcripts responsible for cell development in photoreceptors of the eye and photo-transduction were observed in the Skate population collected from the much shallower sGSL (Lighten et al., 2016).
The above-mentioned mechanisms are believed to be strongly influenced by epigenetics in the case of vertebrates (Lighten et al., 2016). This, in turn, indicates that in an environment with shallower water, there is a decreased necessity for highly light-sensitive eyes, thus justifying the phenotypic adaptation of the sGSL population (Cvekl & Mitton, 2010).
The study concludes with the finding that epigenetic regulation-mediated phenotypic, physiological, and life history-based changes have enabled the species in question to react to environmental challenges in a faster and more efficient manner (Lighten et al., 2016).
Epigenetic adaptations of the three-spined stickleback - are epigenetic adaptations enough?
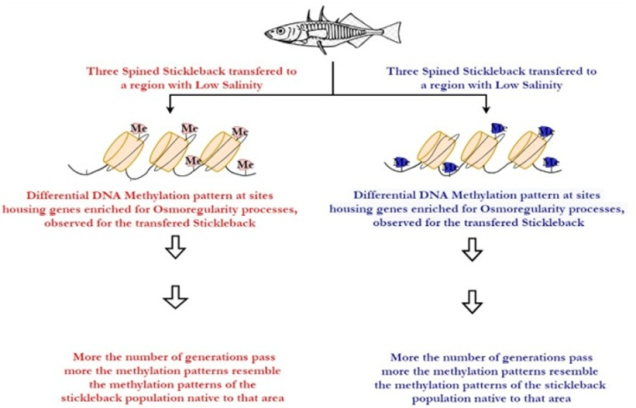
Reusch et al. and Eizaguirre et al. studied the adaptations in the Baltic three-spined stickleback and similar to the previous cases, they observed that certain epigenetic modifications in the three-spined stickleback allow it to survive in a host of salinity conditions (Heckwolf et al., 2020). This is thus another case where epigenetic modification gives rise to ‘phenotypic plasticity’ allowing the organism to adapt to the change in its environment which is brought about by climate change (Heckwolf et al., 2020) (Figure 4).
They also observed that these genetic changes were transgenerational and were transmitted to the offspring as well (Heckwolf et al., 2020). This in turn allowed the offspring to better respond to environmental change (Heckwolf et al., 2020). The improvement of the response of the offspring however was observed to be lesser than anticipated (Heckwolf et al., 2020). The study went on to show that although the epigenetic modifications did indeed confer a certain degree of adaptability to the stickleback, the organism would eventually be unable to keep up with the changing climate in the long term (Heckwolf et al., 2020) (Figure 4). This gives rise to the idea that in the grand scheme of things, epigenetic modifications may be insufficient to keep up with the changing climate (Heckwolf et al., 2020).
|
|
|
Figure 4. An Experiment showing how Transgenerational Epigenetic modifications in the three-spined stickleback gives riseto phenotypic plasticity in the form of salinity tolerance (Heckwolf et al., 2020) |
Climate-change-mediated maladaptive epigenetic changes
Different organisms in both terrestrial as well aquatic habitats can keep up with the changing environment as a result of epigenetic alterations in response to the said changes. Such changes in the genome of the organism, and the resulting phenotypic changes which ensue, are crucial for the survival of the organism. That said, all phenotypes which arise due to these epigenetic modifications need not always be adaptive.
Epigenetic adaptations and rise in invasive species
Epigenetic adaptations allow the organisms to tolerate different changes in the environment due to ‘phenotypic plasticity’ (Carneiro & Lyko, 2020). This ‘phenotypic plasticity’ allows the organism to respond to different changes, even allowing the organisms to thrive in different regions altogether (Carneiro & Lyko, 2020). Such an ability not only greatly reduces the chances of extinction, but also may direct it to colonize different regions where its growth cannot be controlled due to a lack of a natural predator (Campoy et al., 2022). Thus, the organism becomes invasive. Such invasive species are a great threat to biodiversity due to their ability to outcompete the native inhabitants and lead to their extinction.
In plants, epigenetic adaptations allow them to survive in different habitats and environments. A study conducted on Carpobrotus edulis, a perennial plant found in South Africa and invasive in many coastal regions, showed differences in DNA methylation between invasive and native plants (Campoy et al., 2022). These differences result in different phenotypic variations, such as changes in water uptake between the invasive and native plants (Campoy et al., 2022). These changes allow the invasive forms to more efficiently utilize the resources of their invaded habitat, and thereby outcompete the other native plants (Campoy et al., 2022). The authors also showed that alterations in temperature and rainfall result in changes in methylation patterns, and affect nitrogen partitioning in the plants (Campoy et al., 2022).
In animals, different studies have correlated the increased fitness of the invasive species with that of the altered DNA methylation patterns (Ardura et al., 2017; Sheldon et al., 2018). The subjects of said studies range from the invasive house sparrow (Passer domesticus) to the pygmy mussel (Xenostrobus secures), where hypomethylation was implicated as the cause for the expansion of the invasive species (Ardura et al., 2017; Sheldon et al., 2018).
|
|
|
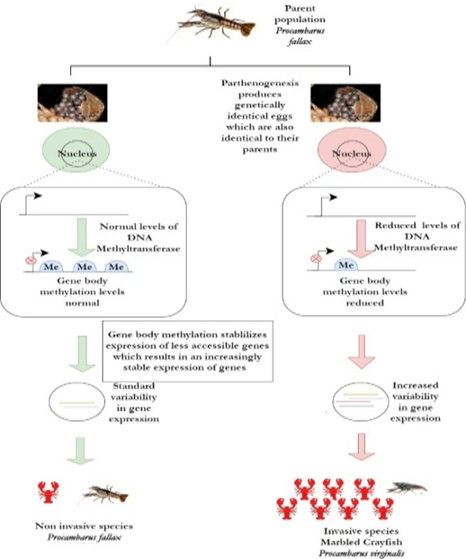
Figure 5. Climate change-mediated epigenetic modifications on crayfish promoting invasiveness (Gatzmann et al., 2018) |
The marbled crayfish (Procambarusvirginalis) is a freshwater crayfish that has invaded different distinct habitats in Madagascar (owing to anthropogenic releases). What is interesting about this species and its role as an invader is that the marbled crayfish undergoes obligatory parthenogenesis; a rare event often described as an “evolutionary scandal” (Gutekunst et al., 2018). As parthenogenesis is a process by which the egg develops into an embryo in the absence of fertilization, the offspring produced are genetically identical to each other as well as the parent (Gutekunst et al., 2018). This severely restricts the genetic diversity of marbled crayfish. Genetic diversity i.e. the presence of different and suitable genes is the strong suit of most invasive species and it is what allows them to colonize these different habitats in the first place. So how does Procambarusvirginaliseven with its restricted genetic diversity invade and colonize different environments? The answer was uncovered by Gatzmann et al. when they studied the DNA methylation patterns in the marbled crayfish (Figure 5). They found that Procambarus virginalis showed reduced gene body methylation when compared to the parent population (P. fallax) (Gatzmann et al., 2018; Genc et al., 2022). This reduced gene body methylation results in increased variability in gene expression which promotes invasiveness (Gatzmann et al., 2018).
Impact of climate change-mediated epigenetic modifications on insect polyphenism
Polyphenism is a remarkable form of ‘phenotypic plasticity’ and is defined as “an individual’s ability to develop into two or more discrete phenotypes” (Richard et al., 2019). Insects can manifest distinct phenotypes based on the environmental stimulus they receive (Richard et al., 2019). Caste-based division of labor amongst honey bees and ant colonies are examples of such polyphenisms.
|
|
|
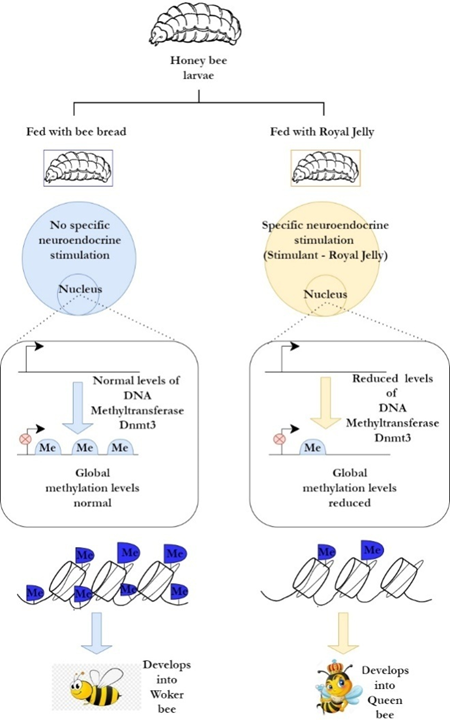
Figure 6. Epigenetic modifications in Honey bee in response to an environmental stimulus (Colton, 1903) |
The environmental stimulus is transmitted to the insects, generally by the neuro-endocrine system, and results in epigenetic modifications which give rise to distinct phenotypes (Richard et al., 2019). For example, in honey bees, consumption of royal jelly leads to decreased Dnmt3 (methyl transferase) expression, resulting in a reduction in global methylation of the genome, allowing the larvae to develop into a queen (Richard et al., 2019). The manifestation of the queen phenotype is opposed to that of a worker phenotype (Figure 6) (Bonasio et al., 2012). This implies that the environment plays a crucial role in determining the composition of different insect populations which is necessary for their survival (Richard et al., 2019).
Alteration in environmental cues mediated by climate change results in altered distribution of the distinct phenotypes in a particular insect population and endangers them (Richard et al., 2019). The light bush brown butterfly Bicylusdorotheafound in West and Central Africa shows polyphenisms in response to seasonal variations (Prudic et al., 2015). These manifest themselves as both physical and behavioral changes (Prudic et al., 2015). One such prominent change is the change in wing spots, which presumably plays an important role in predator escape (Prudic et al., 2015). The dry season which is marked by low food resources results in butterflies with small “cryptic” wingspots (Prudic et al., 2015). Meanwhile, the wet season which is marked by a high abundance of food results in prominent wingspots (Prudic et al., 2015). However, an increase in dryness during the wet season due to climate may lead to the alteration of this pattern hampering the butterfly’s ability to avoid predators (Prudic et al., 2015).
Epigenetic trap - a response to climate change
Different species possess the ability to manipulate their reproductive schemes in terms of the growing alterations in the environment via sex-determination strategies controlled epigenetically (Consuegra & Rodríguez López, 2016). This helps the organisms to adjust themselves to the environment in the face of climate change, and at the same time build up a certain degree of ‘phenotypic plasticity’ (Consuegra & Rodríguez López, 2016). This is possible because the organisms possess the ability to carry on a certain plastic or non-genetic memory of the immediate environment that their parents resided in (Consuegra & Rodríguez López, 2016).
Unfortunately, all epigenetic mechanisms do not offer an adaptive advantage to the organisms under question; rather some of them can turn out to be quite maladjusted or dysfunctional (Consuegra & Rodríguez López, 2016; Alhussain et al., 2022).
Although in certain cases adaptation can be accelerated by epigenetic modifications, it can result in a decline in the overall health of the species (Consuegra & Rodríguez López, 2016). This might occur as a result of what is referred to as the ‘epigenetic trap’, where alterations at the epigenetic level and its subsequent effect on environment-mediated sex determination can enhance the viability or life span of one sex, at the cost of reducing the fitness of the other sex, due to initial alterations in the sex-ratios (Consuegra & Rodríguez López, 2016). This can ultimately disintegrate the original population of a species (Consuegra & Rodríguez López, 2016). For example, in the case of Lamprotornissuperbus or the Superb Starling which resides in erratic environments, it was found that environmental conditions that persisted ahead of egg laying can influence the DNA methylation levels of the promoter of a crucial gene located on the HPA (hypothalamic-pituitary-adrenal) axis responsible for handling stress response (Rubenstein et al., 2016). This in turn offered a survival advantage to the male birds which were born following a bitter change in environmental conditions (Rubenstein et al., 2016). This can be maladaptive, if the environment to which the offspring is being adapted under the influence of the mother, is different from the environment which is prevalent once the offspring hatches (Rubenstein et al., 2016). This is due to the rapid environmental changes which occur due to climate change.
Pollution-mediated epigenetic modifications
DNA methylation is an epigenetic phenomenon that undergoes modifications throughout the lifespan of an individual, spanning the periods of pregnancy through to the declining years, when subjected to air pollution (Rider & Carlsten, 2019). Studies have revealed that the repercussions of DNA methylation are clearer in adulthood, possibly due to extended sessions of exposure to air pollution (Rider & Carlsten, 2019). This suggests that these epigenetic changes need time to accumulate to manifest themselves (Rider & Carlsten, 2019). Interestingly, researchers have also been able to associate alterations in DNA methylation with the occurrence of lung diseases (Rider & Carlsten, 2019). Thus, a better idea as to how pollution can bring about such epigenetic changes might help design strategies to minimize episodes of such diseases (Rider & Carlsten, 2019). Air pollution results in the alteration of the balance between cytosine and 5-methylcytosine, along with global alterations of DNA methylation patterns, resulting in the hypomethylation of promoters of genes encoding for proteins of immunological importance and inflammation (Rider & Carlsten, 2019).
|
|
|
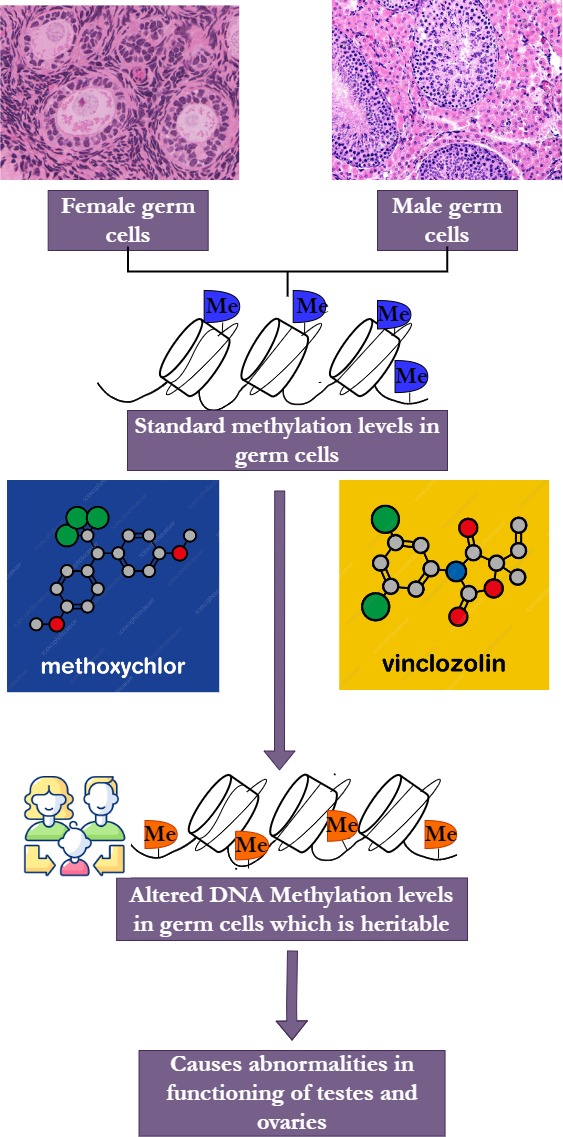
Figure 7. Effect of ChemicalPollutionOn DNA Methylation in Humans (Anway & Skinner, 2006; Guerrero-Bosagna et al., 2010). |
Exposure to chemical pesticides harboring environmental pollutants such as methoxychlor and vinclozolin results in the alteration of DNA methylation levels, which might be inherited (Anway & Skinner, 2006; Guerrero-Bosagna et al., 2010). These modifications occur at the germline level and can result in abnormalities in the functioning of testes or ovaries (Ranjan & Sharma, 2016) (Figure 7).
CONCLUSION
Climate change has turned out to be one of the biggest challenges ever. It is a threat not only to humans but to all living things. Epigenetic modification is a strategy employed by all organisms to combat the rapid changes in the environment and survive.
Pre-existing epigenetic mechanisms allow organisms to rapidly adapt to changing environments. The long-term benefits of such pre-existing mechanisms in the face of climate change are limited making “genetic rescue” the better solution.
Yet not all epigenetic modifications brought about by climate change are helpful. Many modifications imposed on the genomes of organisms by the change in climate threaten their very existence. In addition, climate change may also render a previously adaptive epigenetic response into a maladaptive response causing the organism to fall into an “epigenetic trap”.
Thus, epigenetic modifications in the face of climate change can be likened to a double-edged sword which in most cases confers an adaptive advantage but in some cases may also be detrimental to the organisms involved.
ACKNOWLEDGMENTS: The authors express their deepest gratitude to Rev. Fr. Dr. Dominic Savio, S.J., Principal and Rector, St. Xavier’s College (Autonomous), Kolkata, and Dr. Subhankar Tripathi, Principal, Sarsuna College, Kolkata.
CONFLICT OF INTEREST: None
FINANCIAL SUPPORT: None
ETHICS STATEMENT: None
Alhussain, B., Alremthi, A., Emad, Y., Abdulrahim, A., Alanazi, H., & Alotaibi, S. (2022). Quality of root canal fillings and prevalence of apical radiolucencies; A retrospective study. Annals of Dental Specialty, 10(3), 74-78.
Anway, M. D., & Skinner, M. K. (2006). Epigenetic transgenerational actions of endocrine disruptors. Endocrinology, 147(6), s43-s49. doi:10.1210/en.2005-1058
Ardura, A., Zaiko, A., Morán, P., Planes, S., & Garcia-Vazquez, E. (2017). Epigenetic signatures of invasive status in populations of marine invertebrates. Scientific Reports, 7(1), 1-10. doi:10.1038/srep42193
Bautista, N. M., & Crespel, A. (2021). Within-and trans-generational environmental adaptation to climate change: Perspectives and new challenges. Frontiers in Marine Science, 1337. doi:10.3389/fmars.2021.729194
Bonamour, S., Chevin, L. M., Charmantier, A., & Teplitsky, C. (2019). Phenotypic plasticity in response to climate change: The importance of cue variation. Philosophical Transactions of the Royal Society B, 374(1768), 20180178. doi:10.1098/rstb.2018.0178
Bonasio, R., Li, Q., Lian, J., Mutti, N. S., Jin, L., Zhao, H., Zhang, P., Wen, P., Xiang, H., Ding, Y., et al. (2012). Genome-wide and caste-specific DNA methylomes of the ants camponotus floridanus and Harpegnathos saltator. Current Biology, 22(19), 1755-1764. doi:10.1016/j.cub.2012.07.042
Campoy, J. G., Sobral, M., Carro, B., Lema, M., Barreiro, R., &Retuerto, R. (2022). Epigenetic and phenotypic responses to experimental climate change of native and invasive carpobrotus edulis. Frontiers in Plant Science, 13. www.frontiersin.org.https://www.frontiersin.org/articles/10.3389/fpls2022.888391/full
Carneiro, V. C., & Lyko, F. (2020). Rapid epigenetic adaptation in animals and its role in invasiveness. Integrative and Comparative Biology, 60(2), 267-274. doi:10.1093/icb/icaa023
Cheng, S., Yang, H. J., & Swaroop, A. (2013). Epigenetic regulation of rod photoreceptor development. Investigative Ophthalmology & Visual Science, 54(15), 6111. iovs.arvojournals.org. https://iovs.arvojournals.org/article.aspx?articleid2151128
Colton, B. P. (1903). Zoology: Descriptive and practical (Vol. 1). DC Heath & Company. Available from: https://etc.usf.edu/clipart/6800/6847/honey-bee_6847.htm
Consuegra, S., & Rodríguez López, C. M. (2016). Epigenetic‐induced alterations in sex‐ratios in response to climate change: An epigenetic trap? BioEssays, 38(10), 950-958. doi:10.1002/bies.201600058
Cvekl, A., & Mitton, K. P. (2010). Epigenetic regulatory mechanisms in vertebrate eye development and disease. Heredity, 105(1), 135-151. www.ncbi.nlm.nih.gov. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC228956/
Dixon, G., Liao, Y., Bay, L. K., & Matz, M. V. (2018). Role of gene body methylation in acclimatization and adaptation in a basal metazoan. Proceedings of the National Academy of Sciences, 115(52), 13342-13346. doi:10.1073/pnas.183749115
Eoh, H., & Rhee, K. Y. (2013). Multifunctional essentiality of succinate metabolism in adaptation to hypoxia in Mycobacterium tuberculosis. Proceedings of the National Academy of Sciences, 110(16), 6554-6559. doi:10.1073/pnas.1219375110
Gatzmann, F., Falckenhayn, C., Gutekunst, J., Hanna, K., Raddatz, G., Carneiro, V. C., & Lyko, F. (2018). The methylome of the marbled crayfish links gene body methylation to stable expression of poorly accessible genes. Epigenetics & Chromatin, 11(1), 1-12. doi:10.1186/s13072-018-0229-6
Genc, A., Isler, S. C., Oge, A. E., & Matur, Z. (2022). Effect of sagittal split osteotomy with medpor® porous polyethylene implant on masticatory reflex. Annals of Dental Specialty, 10(3), 12-16.
Guerrero-Bosagna, C., Settles, M., Lucker, B., & Skinner, M. K. (2010). Epigenetic transgenerational actions of vinclozolin on promoter regions of the sperm epigenome. PloS One, 5(9), e13100. doi:10.1371/journal.pone.0013100
Gutekunst, J., Andriantsoa, R., Falckenhayn, C., Hanna, K., Stein, W., Rasamy, J., & Lyko, F. (2018). Clonal genome evolution and rapid invasive spread of the marbled crayfish. Nature Ecology & Evolution, 2(3), 567-573. doi:10.1038/s41559-018-0467-9
Heckwolf, M. J., Meyer, B. S., Häsler, R., Höppner, M. P., Eizaguirre, C., & Reusch, T. B. (2020). Two different epigenetic information channels in wild three-spined sticklebacks are involved in salinity adaptation. Science Advances, 6(12), eaaz1138. doi:10.1126/sciadv.aaz1138
Kingsolver, J. G., & Buckley, L. B. (2017). Evolution of plasticity and adaptive responses to climate change along climate gradients. Proceedings of the Royal Society B: Biological Sciences, 284(1860), 20170386. doi:10.1098/rspb.2017.0386
Lafon-Placette, C., Le Gac, A. L., Chauveau, D., Segura, V., Delaunay, A., Lesage-Descauses, M. C., Hummel, I., Cohen, D., Jesson, B., Le Thiec, D., et al. (2018). Changes in the epigenome and transcriptome of the poplar shoot apical meristem in response to water availability affect preferentially hormone pathways. Journal of Experimental Botany, 69(3), 537-551. doi:10.1093/jxb/erx409
Le Gac, A. L., Lafon-Placette, C., Chauveau, D., Segura, V., Delaunay, A., Fichot, R., Marron, N., Le Jan, I., Berthelot, A., Bodineau, G., et al. (2018). Winter-dormant shoot apical meristem in poplar trees shows environmental epigenetic memory. Journal of Experimental Botany, 69(20), 4821-4837. doi:10.1093/jxb/ery271
Liew, Y. J., Zoccola, D., Li, Y., Tambutté, E., Venn, A. A., Michell, C. T., Cui, G., Deutekom, E. S., Kaandorp, J. A., Voolstra, C. R., et al. (2018). Epigenome-associated phenotypic acclimatization to ocean acidification in a reef-building coral. Science Advances, 4(6), eaar8028. doi:10.1126/sciadv.aar8028
Lighten, J., Incarnato, D., Ward, B. J., van Oosterhout, C., Bradbury, I., Hanson, M., & Bentzen, P. (2016). Adaptive phenotypic response to climate enabled by epigenetics in a K-strategy species, the fish Leucoraja ocellata (Rajidae). Royal Society Open Science, 3(10), 160299. doi:10.1098/rsos.160299
Morgan, R. (2019). The organism-environment interface: How epigenetics can help us understand the impacts of climate change. Retrieved August 10, 2022, from: https://sph.umich.edu/pursuit/219posts/organim environment-interface-how epigenetics-helps-us understand climate-change-impacts.html
Müller, M., & Gailing, O. (2019). Abiotic genetic adaptation in the Fagaceae. Plant Biology, 21(5), 783-795. doi:10.1111/plb.13008
Prudic, K. L., Stoehr, A. M., Wasik, B. R., & Monteiro, A. (2015). Eyespots deflect predator attack increasing fitness and promoting the evolution of phenotypic plasticity. Proceedings of the Royal Society B: Biological Sciences, 282(1798), 20141531.
Ranjan, S., & Sharma, P. K. (2016). Effect of environmental chemical exposures on epigenetics of diseases: Asystematic review. International Journal of Life Sciences Scientific Research, 2(4). doi:10.21276/ijlssr.2016.2.4.11
Rezapour-Nasrabad, R., & Tayyar-Iravanlou, F. (2022). Hippotherapy and its effect on behavioral and executive disorders in children with autism spectrum disorder. Journal of Advanced Pharmacy Education and Research, 12(3), 15-20.
Richard, G., Le Trionnaire, G., Danchin, E., & Sentis, A. (2019). Epigenetics and insect polyphenism: Mechanisms and climate change impacts. Current Opinion in Insect Science, 35, 138-145. doi:10.1016/j.cois.2019.06.013
Rider, C. F., & Carlsten, C. (2019). Air pollution and DNA methylation: Effects of exposure in humans. Clinical Epigenetics, 11(1), 1-15. doi:10.1186/s13148.019-0713-2
Rubenstein, D. R., Skolnik, H., Berrio, A., Champagne, F. A., Phelps, S., & Solomon, J. (2016). Sex‐specific fitness effects of unpredictable early life conditions are associated with DNA methylation in the avian glucocorticoid receptor. Molecular Ecology, 25(8), 1714-1728. doi:10.1111/mec.13483
Sheldon, E. L., Schrey, A., Andrew, S. C., Ragsdale, A., & Griffith, S. C. (2018). Epigenetic and genetic variation among three separate introductions of the house sparrow (Passer domesticus) into Australia. Royal Society Open Science, 5(4), 172185. doi:10.1098/rsos.172185
Sow, M. D., Allona, I., Ambroise, C., Conde, D., Fichot, R., Gribkova, S., Jorge, V., Le-Provost, G., Pâques, L., Plomion, C., et al. (2018). Epigenetics in forest trees: State of the art and potential implications for breeding and management in a context of climate change. Advances in Botanical Research, 88, 387-453. doi:10.1016/bs.abr.2018.09.003
Thiebaut, F., Hemerly, A. S., & Ferreira, P. C. G. (2019). A role for epigenetic regulation in the adaptation and stress responses of non-model plants. Frontiers in Plant Science, 10, 246. doi:10.3389/fpls.2019.00246
United Nations. (n.d.). What is climate change? United Nations. Retrieved October 6, 2022, from: https://www.un.org/en/climatechange/what-is-climate-change
Weyrich, A., Lenz, D., & Fickel, J. (2018). Environmental change-dependent inherited epigenetic response. Genes, 10(1), 4. doi:10.3390/genes10010004
 This work is licensed under a Creative Commons Attribution 4.0 International License.
This work is licensed under a Creative Commons Attribution 4.0 International License.