The genus Pantoea is Gram-negative bacteria, distributed in contaminated soil but the increased numbers of Pantoea species were isolated from human bodies, indicating that these species are opportunistic pathogen and cause nosocomial infections. This research aimed to isolate, characterize, and biocontrol of Pantoea species, isolated and collected from soil. During searching for phosphate-degrading bacteria, isolate MO11 was obtained from soil samples and grow well on a medium containing tricalcium phosphate. It was identified according to morphology, physiology, and molecular methods. The antimicrobial susceptibility patterns to antibiotics were determined and the antibiotic resistance pattern reflects its clinical importance. It showed high resistance to Amoxicillin, Amikacin, and Gentamicin while it was sensitive to Imepinem and Meropenem. The isolate was identified as Pantoea calida with 97% and as P. agglomerans with 95% similarity level. Therefore, it is the first time to isolate this bacterium from the soil in Saudi Arabia. Streptomyces coeruleorubidus MM5 was previously isolated from contaminated soil and it was identified using molecular methods. It showed good inhibitory activity against Pantoea calida using the cross streak method, Agar plug diffusion method, and Agar well diffusion methods. Further, the active agent was extracted by ethyl acetate solvent from the extracellular filtrate and it showed excellent antibacterial activity against Pantoea cells which showed resistance to the routinely used antibiotics. In conclusion, although multidrug-resistant Pantoea cells pose a serious threat worldwide and the appropriate antibiotic must be prescribed after determining the sensitivity test, Streptomyces species have played a critical role as biocontrol agents.
INTRODUCTION
The genus Pantoea is ubiquitous, it is well known for more than ten years as the genus belonged to Enterobacteriaceae and it resembles species of the genera Erwinia and Enterobacter (Akshita et al., 2020; Mirza et al., 2020; Suhartati et al., 2021). Pantoea is facultative anaerobic Gram-negative bacilli with peritrichous flagella (AbdAlhussen & Darweesh, 2016; Sadovnikova et al., 2022). The cultural and biochemical characters and phylogenetic approach are still mainly used to identify various Pantoea spp. Some species of the genus Pantoea have beneficial effects as a producer of antibiotic like pantocin A and B while others infect some plants, but the diversity of Pantoea strains and their possible association with hosts and disease are still to be demonstrated (Cruz et al., 2007; Walterson & Stavrinides, 2015). Its role in phosphorus solubilization which is an important macronutrient, needed for all plant growth is well known. Thus, the solubilization of organic and inorganic insoluble soil phosphate by the eco-friendly bacterium is necessary for agricultural uses (Hegyi et al., 2021; Madrid-Delgado et al., 2021; Zhang et al., 2022).
Pantoea species have been isolated from soil, water, eucalyptus leaves (as epiphytes or endophytes bacteria) and plant seeds and fruits, and, four Pantoea species were isolated and showing symptoms of blight and die-back on the plant shoot while Pantoea allii was recovered from onion leaf and seed (De Maayer et al., 2014; Sheibani-Tezerji et al., 2015; Achbani et al., 2016; Ma et al., 2016; Palmer et al., 2016). Other members of the genus Pantoea have been isolated from animals and human wounds and may be opportunistic pathogens. Some species were isolated from human and animal gastrointestinal tracts, feces, dairy products, human blood, and urine. Also, three new Pantoea species were isolated from infant formula. They cause some human infections like septic arthritis and osteomyelitis. They can live as opportunistic pathogens in the immunocompromised human, causing a wound, blood, and urinary tract infections (Brady et al., 2010). The species of the genus Pantoea included P. agglomerans, P. ananatis, P. citrea, P. dispersa, P. punctata, P. stewartii, and P. terrea were isolated and identified (Schmid et al., 2003). In recent years, a variety of human diseases due to Pantoea species are increased and nosocomial infections by this organism were reported in different hospital intensive care units, burn, hemodialysis, and oncology unit. Pantoea spp. was isolated during an outbreak in a Malaysian neonatal intensive care unit in 2006 (Van Rostenberghe et al., 2006; Liberto et al., 2009). In India, Tiwari and Beriha (2015) isolated P. agglomerans from neonatal sepsis and it is difficult to treat this bacterium. In Iraq, different antibiotics resistance Pantoea spp were isolated from different clinical and environmental sources (AbdAlhussen & Darweesh, 2016).
During the past decade, increasing antibiotic resistance to currently used drugs cause high morbidity and mortality throughout the world, and novel alternative antibiotics with the heterocyclic structure are needed. Actinomycetes, the filamentous Gram-positive, aerobic spore-forming bacteria had DNA rich in GC content (60-80 %) and is the most attractive bacteria with abilities to produce unique or novel bioactive compounds, beneficial to humans against some pathogens like multidrug-resistant bacteria, Serratia marcescens, Staph aureus and MRSA (Khamna et al., 2010; Genilloud, 2017; Kemung et al., 2018; Takahashi & Nakashima, 2018). The soil had microbial diversity of Actinomycetes which form extensive branching substrate and aerial mycelia and play significant geochemical roles in soil. They are highly valued for their capacity to form biological secondary metabolites with antimicrobial activities and today, most of the used antibiotics are Actinomycetes natural secondary products (Barton, 2006). On another hand, unexplored soil areas have a significant potential for exploring new bacterial isolates that present under increased competition in the environment like limited nutrient content and producing antimicrobial substances against each other. Some Streptomyces isolates produce many secondary metabolites with excellent inhibitory activities against different human microbial pathogens (Sadoway et al., 2013; Al-Ansari et al., 2019). Production of bioactive agents or antibiotics by actinobacteria with antibacterial and/or antitumor by members of the genus Streptomyces, Amycolatopsis, Micromonospora, and Nocardia were documented (Nakaew et al., 2009; Arifuzzaman et al., 2010; Rangseekaew & Pathomaree, 2019; Nakagawa et al., 2022). Out of 22,000 bioactive secondary metabolites, 16,000 compounds were selected as bioactive antibiotic activities and about 45% of these compounds were from Actinomycetes, and more than 8000 useful bioactive compounds were from the genus Streptomycetes (Berdy, 2005; Khamna et al., 2010). From soil, S. coelicolor and S. violaceoruber produce an important antibiotic actinorhodin with a wide range of applications in the medical sector (Palanichamy et al., 2001). Streptomyces sp. SM01 was isolated from Indian soil and it produced a novel antibiotic picolinamycin which was effective against multi-drug resistant bacteria (Maiti et al., 2020). The present study was focused on the isolation and identification of Pantoea species from soil and its biocontrol by soil actinomycetes.
MATERIALS AND METHODS
Collection of soil samples
Soil samples were collected in sterilized bags from different depths, 5, 15, and 20 cm, from contaminated soil, collected from Jeddah, Saudi Arabia. Soil samples were kept under 4ºC until used for bacterial isolation on nutrient agar using serial dilutions methods. The obtained bacterial colonies were selected, purified, and preserved on the same medium.
Phosphate solubilization and indole production
All the bacterial isolates were screened for phosphate solubilization on phosphate mineral agar containing g/l: 10 glucose, 5 Ca3(PO4)2, 5 MgCl2.6 H2O, 0.25, MgSO4 7H2O, 0.2 KCl, 0.1 (NH4)2SO4 (Nautiyal, 1999) and all plates were incubated for 5 days at 30°C. The colonies with clear halo zones were considered to be phosphate-solubilizing bacteria and the mean diameter of the clear zone (mm) around the tested bacterial colony was measured (Janardan & Verma, 2012). The bacterial growth was measured and represented as +++ (high growth) and ++ (moderate growth). The positive bacteria with clear zone were selected for indole acetic acid production (IAA) in a medium supplemented with 2 mg/ml of L-tryptophan and IAA was extracted from the culture filtrate with ethyl acetate and the quantity was detected according to the method described before by measuring the A530 nm and preparing a standard curve of IAA (Bano & Musarrat, 2003; Ahmed et al., 2004). IAA-producing Azotobacter sp. was obtained from the culture collection of the faculty of Science, KAU, Jeddah, Saudi Arabia, and used as the reference strain.
The used actinomycete isolate
The isolate Streptomyces coeruleorubidus MM5 was previously isolated from the industrial zone, Jeddah (Aburas et al., 2021) and kindly provided from the culture collection of the faculty of Science, KAU, Jeddah, Saudi Arabia. It was grown on a Starch Nitrate agar medium (Shirling & Gottlieb, 1966), examined under light and scanning microscopes at the KHMR center, KAU, and preserved on the same medium.
Preparation of the bacterial suspension
The bacterial suspension was prepared in a sterile Falcon tube containing 3 ml of sterile nutrient broth medium, these tubes were incubated at 30˚C for 24 hrs and growth was measured at OD 520 nm using Spectrophotometer and it was adjusted to 0.5.
Screening of Actinomycetes for antimicrobial activity
The Actinomycete isolate, Streptomyces coeruleorubidus MM5 was screened for inhibitory substances production against the tested bacterium MO11 using different methods. Primary screening using Cross streak method is applied by inoculating a single streak of the selected isolate on the center of the Mueller Hinton agar plate and incubating for 5 days at 30ºC, then the plates are seeded with the isolate MM11 in a perpendicular arrangement and the plates were re-incubated again for 24 hrs (Oskay, 2009). Also, the Agar plug diffusion method was used to determine the inhibitory activity of the selected actinomycete against the isolate MO11. By sterile cork borer, a disc of 8 mm diameter of bacterial growth was put on the center of the Mueller Hinton agar plate inoculated with the tested pathogen MO11. The antimicrobial activity of the bacterial growth disc was detected by the inhibition zone (mm). Also, the selected actinomycete, S. coeruleorubidus MM5 was grown on Starch Nitrate agar medium at 30˚C for 5 days at 80 rpm and the culture supernatant was filtrated sterilized through bacterial filter 0.22 µm and the filtrate was extracted with ethyl acetate (1:1V/V). The extract was dried and dissolved in DMSO. Then, 100 µl of the extract was used to fill each agar well with dimmer 6 mm on Muller Hinton agar plates or blood agar inculcated by the tested bacterium, and the plates were incubated at 30˚C for 24 hrs. The mean diameter of the produced inhibition zone was determined (Magaldi et al., 2004; Valgas et al., 2007; Aljamal, 2022).
Characterization of the selected bacterium
The morphological characterization and biochemical tests were carried out for the most active bacterial isolate MO11. The selected isolate was examined on Nutrient, MacConkey, and blood agar plates after incubation at 30˚C under aerobic conditions for 2 days. Cells were examined under a light microscope using an oil immersion lens after Gram staining. Catalase test, oxidase test, starch hydrolysis, coagulate test, blood hemolytic, and antibiotic sensitivity were also recorded. VITEK-2 system at KAUH was used for bacterial identification. Antibiotic susceptibility testing was carried out on Mueller-Hinton agar by "The Kirby-Bauer disc diffusion method. The fresh bacterial suspension was prepared with OD 0.5 and inoculated on Mueller-Hinton plates and antibiotic discs were applied o the agar surface. All the inoculated plates were incubated at 30◦C for 24 hrs, and the susceptibility of the tested isolate to each antimicrobial agent was recorded.
Molecular identification of selected isolate
The selected isolate MO11 was grown on Nutrient agar medium for 2 days at 30˚C, then, DNA was extracted, purified, and amplified using twp primers. After PCR, the sequences of the 16S rRNA were determined at Macrogen. The information on the used primer was summarized in Table 1.
Table 1. Primer used for molecular identification of the isolate MO11
|
The information on the used Primers |
|
|
PCR (primer name and sequence) |
Sequence (primer name and sequence) |
|
27F 5' (AGA GTT TGA TCM TGG CTC AG) 3 |
785F 5' (GGA TTA GAT ACC CTG GTA) 3' |
|
1492R 5' (TAC GGY TAC CTT GTT ACG ACT T) 3' |
907R 5' (CCG TCA ATT CMT TTR AGT TT) 3' |
Statistical analysis
Statistical Package for Social Science (SPSS for windows, version 17) was used for comparing the data, a mean value of three reading ± SD was used and a t-test was used to detect any significant difference between the sample and control. Significant results were obtained at P<0.05.
RESULTS AND DISCUSSION
A bacterium was isolated from soil samples, collected from the contaminated area. Out of 20 bacterial isolates obtained on Nutrient agar, 7 isolates were phosphate-solubilizing bacteria and they had different abilities to produce IAA (Table 2). The isolate MO11 was the most active isolate, thus it was selected for more detailed studies.
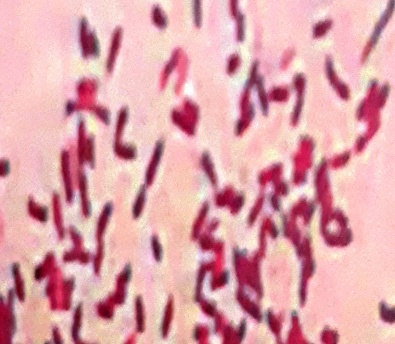
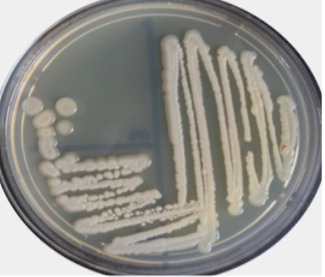
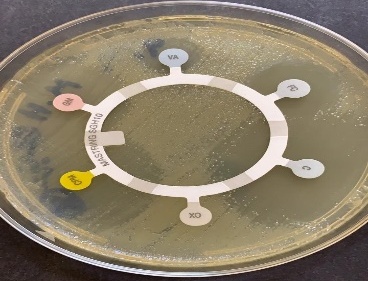
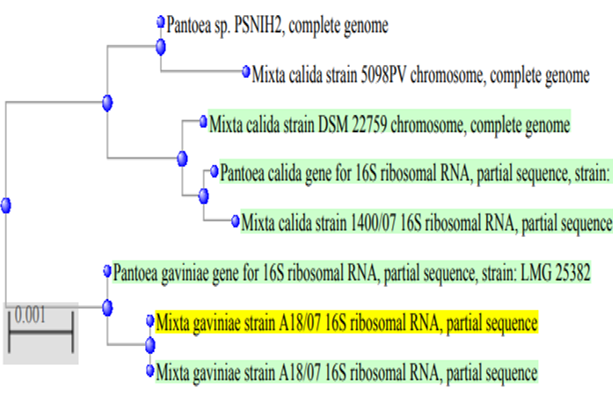
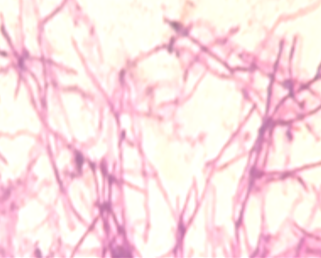
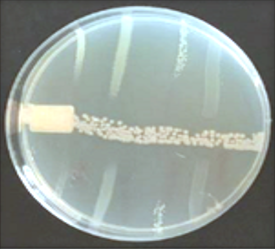
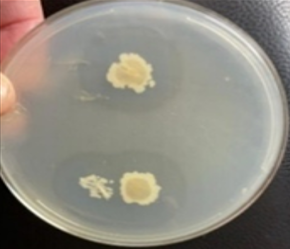
Under light microscopic, the selected isolate MO11 appeared as Gram-negative motile bacilli (Figure 1a). It grows well on different agar media, Nutrient agar (Figure 1b), MacConkey agar, and blood agar plates. It appeared as punctuate circle smooth colonies, middle in size, while on blood agar it produce smooth non-hemolytic colonies. It had positive results for catalase, indole, and citrate production and fermentation of Raffinose. It showed negative results to produce H2S and other biochemical reactions had variable results (Table 3). The isolate MO11 was resistant to Amoxicillin, Amikacin, and Gentamicin while it was sensitive to Imepinem and Meropenem (Figure 1c). The isolate MO11 was identified by an automated VITEK-2 compact system using GN-ID cards as Pantoea spp. A phylogenic analysis was performed to identify the selected isolate MO11using a partial sequence of the 16S rRNA which showed a high level of sequence similarity with member genus Pantoea with a homology level of 97% to P. calida as in the phylogenic analysis (Figure 2).
Table 2. The growth of the obtained bacterial isolates on nutrient agar, Gram strain, production of IAA, and phosphate solubilization
|
Isolate |
Soil depth (cm) |
Shape |
Color |
Gram stain |
IAA (mg/l) |
Phosphate solubilization |
|
|
Growth |
Halo Zone Diameter (mm) |
||||||
|
MM1 |
5 cm |
Monococcus |
White |
G +ve |
1.19±0.12* |
++ |
14±1.9* |
|
MM4 |
5 cm |
Bacilli |
Yellow |
G -ve |
ND |
++ |
12±1.3* |
|
MM9 |
1 5 cm |
Bacilli |
White |
G + |
1.45±0.14* |
++ |
18±2.1* |
|
MM11 |
15 cm |
Bacilli |
White |
G- ve |
0.49±0.49* |
+++ |
34±2.0 |
|
MM15 |
20 cm |
diplococci |
White |
G -ve |
1.69±0.11* |
++ |
22±2.8* |
|
MM16 |
5 cm |
Bacilli |
Yellow |
G-ve |
2.89±0.22 |
++ |
21±1.0* |
|
MM20 |
5 cm |
Bacilli |
Gray |
G-ve |
1.90±0.09* |
++ |
20±1.9* |
|
Control (Azotobacter sp.) |
Cocci |
Black |
G-ve |
3.11 |
++ |
30±5.0 |
|
|
+++: high growth, ++: moderate growth, IAA: Indole acetic acid, G+ve: gram-positive, G-ve: gram-negative, ND: Not detected, *: Significant differences compared to control. |
|||||||
Table 3. Biochemical test for identification of Pantoea sp.MO11
|
Test |
Result |
Test |
Result |
|
Shape |
Small rod |
Citrate |
+ |
|
Catalase |
+ |
VP |
+ |
|
Oxidase |
- |
Gas |
+ |
|
Gelatinase |
- |
H2S |
- |
|
Hemolysis |
γ |
Urease |
+ |
|
Motility |
+ |
Indole |
+ |
|
Fermentation of Raffinose |
+ |
MR |
- |
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
Figure 1. The isolate MO11, a) After Gram stain, b) grown on nutrient agar, c) resistance of the isolate to some antibiotics. |
|
|
|
Figure 2. The phylogenic tree of the isolate MO11 and the most related isolates |
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
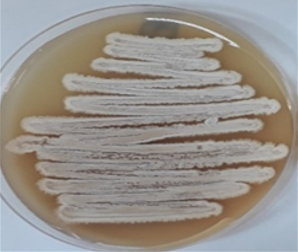
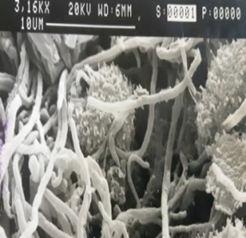
Figure 3. Streptomyces coeruleorubidus MM5, grow on starch nitrate agar (a) for 7 days, under a scanning electron microscope (b) and after Gram stain (c). |
|
|
|
|
a) |
b) |
|
|
|
|
c) |
d) |
|
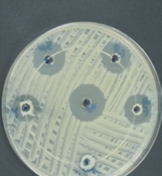
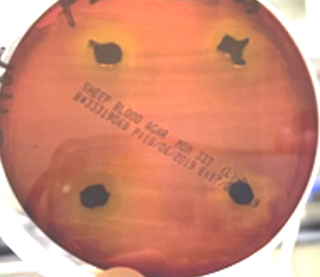
Figure 4. The inhibitory activity of Streptomyces coeruleorubidus MM5 against Pantoea calida MO11 using the three tested techniques, cross streak method (a), Agar plug diffusion method (b), and Agar well diffusion methods on Muller Hinton (c) and blood agar (d). |
|
Isolate Streptomyces coeruleorubidus MM5 was grown on starch nitrate agar for 5 days and examined under a light and scanning microscope. Streptomyces coeruleorubidus MM5 has a dark yellow color with aerial and substrate well-developed mycelia (Figure 3). It showed good inhibitory activity against Pantoea calida MO11 using the three tested techniques, cross streak method Agar plug diffusion method, and Agar well diffusion method (Figure 4). It is noticed that the ethyl acetate extracts of the isolate S. coeruleorubidus MM5 gave a good inhibition zone using agar well diffusion methods either on Muller Hinton or blood agar.
During the screening of soil bacteria for their ability for phosphate solubilization, many isolates showed excellent activity and they also produce IAA in a liquid medium. Isolate MO11 was the most active isolate and it was identified using morphological and physiological methods (Grimont & Grimont, 2005; De Maayer et al., 2014). After molecular studies, it was identified as a bacterial pathogen of the genus Pantoea using 16S rDNA sequencing which is mainly used as a most powerful technique for bacterial identification (Yokota, 1997; Xu et al., 2021). The 16S rDNA sequence was determined and the phylogenic tree was obtained. Many authors reported that Pantoea species can produce IAA and auxin-related compounds which affect the root system structure and shape (Duca et al., 2014; Spaepen, 2015; Stringlis et al., 2018). Pantoea has pigmented a rod-shaped, non-spore-forming, Gram-negative bacterium, belonging to the Enterobacterales order, mainly isolated from soil, urine or associated with plants (Walterson & Stavrinides, 2015; Zhang et al., 2015) and may be used as a growth-promoting bacteria or as biocontrol agents (Dutkiewicz et al., 2016; Luziatelli et al., 2020a, 2020b). It processes an excellent ability to remove heavy metals. In contrast, some species of the genus Pantoea is the obligate infectious agent in humans and can cause opportunistic wound infections or act as hospital-acquired pathogens, mostly in immunocompromised individuals. Septic arthritis, endophthalmitis, periostitis, endocarditis, and osteomyelitis or synovitis in addition to epidemics of nosocomial septicemia were recorded as a frequent clinical result of external infection with P. agglomerans (Dutkiewicz et al., 2016). They added that exposure of hospitalized patient or immunodeficient individuals to medical equipment or fluids contaminated with this bacterium cause serious infection and many fatal cases were recorded in several countries. This bacterium infects cultivated plants, cotton, onion, rice, maize, and sorghum, causing serious diseases or acting as a tumourigenic agent, inducing gall formation on some plant roots. Moreover, it was resistant to most used antibiotics and the resistance increased with time. They are implicated in nosocomial outbreaks of neonates and immunocompromised patients (Habsah et al., 2005). The prevalence and antibiotic susceptibility of Pantoea species during 12 years was recorded by Gajdács (2019) who studied 70 isolates of Pantoea spp. and the highest resistance was to amoxicillin/clavulanic acid and ampicillin, thus the development of multidrug-resistant (MDR) Gram-negative bacteria is a serious concern, and the development of MDR Pantoea spp. may be expected in the future.
The search for novel antibiotics and other bioactive microbial metabolites from soil actinomycetes especially Streptomycetes which for a long time have been recognized as high producers of useful bioactive compounds with many applications is increased. Therefore, it is important to fight against new and emerging pathogens by application of new antimicrobial agents from actinomycetes which form unique natural agents and bioactive metabolites with great interest. Streptomyces species is still a promising and fruitful source to obtain new antibiotics (Berdy, 2005). S. coeruleorubidus have a good ability to produce natural bioactive antimicrobial material which was extracted and showed inhibition activity against Pantoea, thus it could be very useful for pharmaceutical and agricultural uses.
The used S. coeruleorubidus generally showed good growth and inhibited the growth of P. calida using the cross streak method which is one of the best techniques to detect the inhibitor activity of any isolates. This technique is a primary screening method and required a short incubation time to get the result. The isolate showed good inhibitory activity in primary screening, it was subjected to secondary screening using the agar plug method which is often used to highlight the antagonism between microorganisms). S. coeruleorubidus showed high antagonistic activity against P. calida using the plug diffusion method. Moreover, the Agar well diffusion method is widely used to evaluate the antimicrobial activity of plants and microbial extract (Magaldi et al., 2004; Valgas et al., 2007). Ethyl acetate was standardized as the best solvent to extract secondary materials. Similarly, the crude extract of Streptomyces sp. showed high activity against Gram-positive and negative bacteria in addition to MRSA strains. These organisms may produce a high molecular weight glycopeptide with novel anti-MRSA activity (Reddy et al., 2011; Park et al., 2014; Tan et al., 2015).
CONCLUSION
Screening of various actinomycetes for the production of the new drug is a continuous process that should be effective against present-day’s antibiotic-resistant pathogenic bacteria. Actinomycetes have been proven as important sources of various useful secondary metabolites. Also, a recent investigation indicated the potential of marine Streptomyces sp. as a sustainable and useful source of novel bioactive compounds. Hence, the findings of this study revealed that Streptomyces sp. with antibiotic substances production capability was an important application. Also, Streptomyces species from unexplored regions are likely to yield novel antibacterial agents. Streptomyces sp. could be a promising microorganism for the development of a novel antibacterial drug against a wide range of pathogenic bacteria.
ACKNOWLEDGMENTS: None
CONFLICT OF INTEREST: None
FINANCIAL SUPPORT: None
ETHICS STATEMENT: None
AbdAlhussen, L. S., & Darweesh, M. F. (2016). Prevelance and antibiotic susceptibility patterns of Pantoea spp. isolated form clinical and environmental sources in Iraq. International Journal of ChemTech Research, 9(8), 430-437.
Aburas, M., Alzahrani, E. J., & Aly, M. M. (2021). Molecular identification of some filamentous bacteria isolated from contaminated soil for poly hydroxyl butyrate degradation. Bioscience Biotechnology Research Communications, 14(3).
Achbani, E. H., Mazouz, H., Benbouazza, A., & Sadik, S. (2016). First detection of Pantoea ananatis, the causal agent of bacterial center rot of onion in Morocco. Journal of Crop Protection, 5(1), 11-17.
Ahmed, W., Shahroona, B., Zahir, Z. A., & Arshad, M. (2004). Inoculation with ACC-deaminase containing rhizobacteria for improving growth and yield of wheat. Pakistan Journal of Agricultural Sciences, 41(3/4), 119-124.
Akshita, C., Vijay, B. V., & Praveen, D. (2020). Evaluation of phytochemical screening and antimicrobial efficacy of Mesua Ferrea and Piper Cubeba fruit extracts against multidrug-resistant bacteria. Pharmacophore, 11(2), 15-20.
Al-Ansari, M., Alkubaisi, N., Vijayaragavan, P., & Murugan, K. (2019). Antimicrobial potential of streptomyces sp. to the gram positive and gram negative pathogens. Journal of Infection and Public Health, 12(6), 861-866.
Aljamal, J. (2022). Effect of lysine and histidine residues modification on the voltage gating of the mitochondrial porin. Journal of Biochemical Technology, 13(3), 7-12.
Arifuzzaman, M., Khatun, M. R., & Rahman, H. (2010). Isolation and screening of actinomycetes from Sundarbans soil for antibacterial activity. African Journal of Biotechnology, 9(29), 4615-4619.
Bano, N., & Musarrat, J. (2003). Characterization of a new Pseudomonas aeruginosa strain NJ-15 as a potential biocontrol agent. Current Microbiology, 46(5), 0324-0328.
Barton, H. A. (2006). Introduction to cave microbiology: A review for the non-specialist. Journal of Cave and Karst Studies, 68(2), 43-54.
Berdy, J. (2005). Bioactive microbial metabolites. The Journal of Antibiotics, 58(1), 1-26.
Brady, C. L., Cleenwerck, I., Venter, S. N., Engelbeen, K., De Vos, P., & Coutinho, T. A. (2010). Emended description of the genus Pantoea, description of four species from human clinical samples, Pantoea septica sp. nov., Pantoea eucrina sp. nov., Pantoea brenneri sp. nov. and Pantoea conspicua sp. nov., and transfer of Pectobacterium cypripedii (Hori 1911) Brenner et al. 1973 emend. Hauben et al. 1998 to the genus as Pantoea cypripedii comb. nov. International Journal of Systematic and Evolutionary Microbiology, 60(10), 2430-2440.
Cruz, A. T., Cazacu, A. C., & Allen, C. H. (2007). Pantoea agglomerans, a plant pathogen causing human disease. Journal of Clinical Microbiology, 45(6), 1989-1992.
De Maayer, P., Chan, W. Y., Rubagotti, E., Venter, S. N., Toth, I. K., Birch, P. R., & Coutinho, T. A. (2014). Analysis of the Pantoea ananatis pan-genome reveals factors underlying its ability to colonize and interact with plant, insect and vertebrate hosts. BMC Genomics, 15(1), 1-14.
Duca, D., Lorv, J., Patten, C. L., Rose, D., & Glick, B. R. (2014). Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek, 106(1), 85-125.
Dutkiewicz, J., Mackiewicz, B., Lemieszek, M. K., Golec, M., & Milanowski, J. (2016). Pantoea agglomerans: A mysterious bacterium of evil and good. Part IV. Beneficial effects. Annals of Agricultural and Environmental Medicine, 23(2), 197-205.
Gajdács, M. (2019). Epidemiology and antibiotic resistance trends of Pantoea species in a tertiary-care teaching hospital: A 12-year retrospective study. Developments in Health Sciences, 2(3), 72-75.
Genilloud, O. (2017). Actinomycetes: Still a source of novel antibiotics. Natural Product Reports, 34(10), 1203-1232.
Grimont, P. A. D., & Grimont, F. (2005). Genus XXIII. Pantoea. Bergey’s Manual of Systematic Bacteriology, 2, 713-720.
Habsah, H., Zeehaida, M., Van Rostenberghe, H., Noraida, R., Pauzi, W. W., Fatimah, I., Rosliza, A. R., Sharimah, N. N., & Maimunah, H. (2005). An outbreak of pantoea spp. in a neonatal intensive care unit secondary to contaminated parenteral nutrition. Journal of Hospital Infection, 61(3), 213-218.
Hegyi, A., Nguyen, T. B. K., & Posta, K. (2021). Metagenomic analysis of bacterial communities in agricultural soils from Vietnam with special attention to phosphate solubilizing bacteria. Microorganisms, 9(9), 1796.
Janardan, Y., & Verma, J. P. (2012). Isolation and characterization of effective plant growth promoting rhizobacteria from rice rhizosphere of Indian soil. Asian Journal of Biological Sciences, 5(6), 294-303.
Kemung, H. M., Tan, L. T. H., Khan, T. M., Chan, K. G., Pusparajah, P., Goh, B. H., & Lee, L. H. (2018). Streptomyces as a prominent resource of future anti-MRSA drugs. Frontiers in Microbiology, 9, 1-26
Khamna, S., Yokota, A., Peberdy, J. F., & Lumyong, S. (2010). Indole-3-acetic acid production by streptomyces sp. isolated from some Thai medicinal plant rhizosphere soils. EurAsian Journal of BioSciences, 4, 23-32.
Liberto, M. C., Matera, G., Puccio, R., Lo Russo, T., Colosimo, E., & Focà, E. (2009). Six cases of sepsis caused by Pantoea agglomerans in a teaching hospital. The New Microbiologica, 32(1), 119-123.
Luziatelli, F., Ficca, A. G., Bonini, P., Muleo, R., Gatti, L., Meneghini, M., & Ruzzi, M. (2020a). A genetic and metabolomic perspective on the production of indole-3-acetic acid by Pantoea agglomerans and use of their metabolites as biostimulants in plant nurseries. Frontiers in Microbiology, 1475.
Luziatelli, F., Ficca, A. G., Cardarelli, M., Melini, F., Cavalieri, A., & Ruzzi, M. (2020b). Genome sequencing of Pantoea agglomerans C1 provides insights into molecular and genetic mechanisms of plant growth-promotion and tolerance to heavy metals. Microorganisms, 8(2), 153.
Ma, Y., Yin, Y., Rong, C., Chen, S., Liu, Y., Wang, S., & Xu, F. (2016). Pantoea pleuroti sp. nov., isolated from the fruiting bodies of Pleurotus eryngii. Current Microbiology, 72(2), 207-212.
Madrid-Delgado, G., Orozco-Miranda, M., Cruz-Osorio, M., Hernández-Rodríguez, O. A., Rodriguez-Heredia, R., Roa-Huerta, M., & Avila-Quezada, G. D. (2021). Pathways of phosphorus absorption and early signaling between the mycorrhizal fungi and plants. Phyton, 90(5), 1321-1338.
Magaldi, S., Mata-Essayag, S., De Capriles, C. H., Pérez, C., Colella, M. T., Olaizola, C., & Ontiveros, Y. (2004). Well diffusion for antifungal susceptibility testing. International Journal of Infectious Diseases, 8(1), 39-45.
Maiti, P. K., Das, S., Sahoo, P., & Mandal, S. (2020). Streptomyces sp SM01 isolated from Indian soil produces a novel antibiotic picolinamycin effective against multi drug resistant bacterial strains. Scientific Reports, 10(1), 1123-33.
Mirza, A. S., Baig, M. T., Huma, A., Ibrahim, S., Shahid, U., Jabeen, A., & Shaikh, S. (2020). Antibacterial activity of methanol extract of capparis decidua edgew (Forssk.) against staphylococcus aureus, bacillus cereus, salmonella typhi, and escherichia coli. Pharmacophore, 11(4), 46-50.
Nakaew, N., Pathom-Aree, W., & Lumyong, S. (2009). First record of the isolation, identification and biological activity of a new strain of Spirillospora albida from Thai cave soil. Actinomycetologica, 23(1), 1-7.
Nakagawa, N., Odanaka, K., Ohara, H., Ito, T., Kisara, S., & Ito, K. (2022). Effect of smartphone location on pharmacy students’ attention and working memory. Journal of Advanced Pharmacy Education and Research, 12(2), 84-90.
Nautiyal, C. S. (1999). An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiology Letters, 170(1), 265-270.
Oskay, M. (2009). Antifungal and antibacterial compounds from Streptomyces strains. African Journal of Biotechnology, 8(13), 3007-3017.
Palanichamy, V., Hundet, A., Mitra, B., & Reddy, N. (2011). Optimization of cultivation parameters for growth and pigment production by Streptomyces spp. isolated from marine sediment and rhizosphere soil. International Journal of Plant, Animal and Environmental Sciences, 1(3), 158-170.
Palmer, M., De Maayer, P., Poulsen, M., Steenkamp, E. T., Van Zyl, E., Coutinho, T. A., & Venter, S. N. (2016). Draft genome sequences of Pantoea agglomerans and Pantoea vagans isolates associated with termites. Standards in Genomic Sciences, 11(1), 23.
Park, H. B., Lee, J. K., Lee, K. R., & Kwon, H. C. (2014). Angumycinones A and B, two new angucyclic quinones from Streptomyces sp. KMC004 isolated from acidic mine drainage. Tetrahedron Letters, 55(1), 63-66.
Rangseekaew, P., & Pathom-Aree, W. (2019). Cave actinobacteria as producers of bioactive metabolites. Frontiers in Microbiology, 10, 387.
Reddy, N. G., Ramakrishna, D. P. N., & Raja Gopal, S. V. (2011). A morphological, physiological and biochemical studies of marine Streptomyces rochei (MTCC 10109) showing antagonistic activity against selective human pathogenic microorganisms. Asian Journal of Biological Sciences, 4(1), 1-14.
Sadovnikova, N., Lebedinskaya, O., Bezrukov, A., & Davletshina, L. (2022). The correlation between residential property prices and urban quality indicators. Journal of Advanced Pharmacy Education and Research, 12(2), 98-103.
Sadoway, T., Rule, D., Watson, K., Moote, P., Soliman, L.C., Azad, N., Donkor, K., & Horne, D. (2013). Cure from the cave: Volcanic cave actinomycetes and their potential in drug discovery. International Journal of Speleology, 42(1), 35-47.
Schmid, H., Schubert, S., Weber, C., & Bogner, J. R. (2003). Isolation of a Pantoea dispersa-like strain from a 71-year-old woman with acute myeloid leukemia and multiple myeloma. Infection, 31(1), 66-67.
Sheibani-Tezerji, R., Naveed, M., Jehl, M. A., Sessitsch, A., Rattei, T., & Mitter, B. (2015). The genomes of closely related Pantoea ananatis maize seed endophytes having different effects on the host plant differ in secretion system genes and mobile genetic elements. Frontiers in Microbiology, 6, 440.
Shirling, E. T., & Gottlieb, D. (1966). Methods for characterization of streptomyces species. International Journal of Systematic Bacteriology, 16(3), 313-340.
Spaepen, S. (2015). Plant hormones produced by microbes. In Principles of plant-microbe interactions (pp. 247-256). Springer, Cham.
Stringlis, I. A., Zhang, H., Pieterse, C. M., Bolton, M. D., & de Jonge, R. (2018). Microbial small molecules–weapons of plant subversion. Natural Product Reports, 35(5), 410-433.
Suhartati, T., Fatimah, N., Yandri, Y., Kurniawan, R., Bahri, S., & Hadi, S. (2021). The anticancer, antimalarial, and antibacterial activities of moracalkon a isolated from Artocarpus kemando Miq. Journal of Advanced Pharmacy Education & Research, 11(4), 105-110.
Takahashi, Y., & Nakashima, T. (2018). Actinomycetes, an inexhaustible source of naturally occurring antibiotics. Antibiotics, 7(2), 1-2.
Tan, L. T. H., Ser, H. L., Yin, W. F., Chan, K. G., Lee, L. H., & Goh, B. H. (2015). Investigation of antioxidative and anticancer potentials of Streptomyces sp. MUM256 isolated from Malaysia mangrove soil. Frontiers in Microbiology, 6, 1316.
Tiwari, S., & Beriha, S. S. (2015). Pantoea species causing early onset neonatal sepsis: A case report. Journal of Medical Case Reports, 4(9),188.
Valgas, C., Souza, S. M. D., Smânia, E. F., & Smânia Jr, A. (2007). Screening methods to determine antibacterial activity of natural products. Brazilian Journal of Microbiology, 38, 369-380.
Van Rostenberghe, H., Noraida, R., Wan Pauzi, W. I., Habsah, H., Zeehaida, M., Rosliza, A. R., Fatimah, I., Nik Sharimah, N. Y., & Maimunah, H. (2006). The clinical picture of neonatal infection with Pantoea species. Japanese Journal of Infectious Diseases, 59(2), 120-121.
Walterson, A. M., & Stavrinides, J. (2015). Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiology Reviews, 39(6), 968-984.
Xu, N., Qiu, C., Yang, Q., Zhang, Y., Wang, M., Ye, C., & Guo, M. (2021). Analysis of phenol biodegradation in antibiotic and heavy metal resistant Acinetobacter lwoffii NL1. Frontiers in Microbiology, 12, 2670.
Yokota, A. (1997). Phylogenic relationship of actinomyces. Atlas of actinomyces, Asakura Publishing Co. Ltd., Japan, 194-196.
Zhang, C., Cai, K., Li, M., Zheng, J., & Han, Y. (2022). Plant-growth-promoting potential of PGPE isolated from Dactylis glomerata L. Microorganisms, 10(4), 731.
Zhang, Y., Yang, L., & Qiu, Z. (2015). The design strategy of motor vehicle parking lot in colleges and universities—Take the planning and design of huaihai institute of technology as an example. Urban Architecture, 11, 3.
 This work is licensed under a Creative Commons Attribution 4.0 International License.
This work is licensed under a Creative Commons Attribution 4.0 International License.